Abstract
The vitamin D endocrine system in now recognized as subserving a wide range of fundamental biological functions in cell differentiation, inhibition of cell growth as well as immunomodulation. Both forms of immunity, namely adaptive and innate, are regulated by 1,25(OH)2D3. The immune-modulatory properties of vitamin D suggest that it could play a potential therapeutic role in prevention of type 1 diabetes mellitus (T1DM). It is postulated that large doses of vitamin D supplementation may influence the pattern of immune regulation and subsequent progression to T1DM in a genetically susceptible individual. More studies are required to substantiate the relation between T1DM and vitamin D/vitamin D analogues in the pattern of immune regulations in susceptible individuals. In type 2 diabetes mellitus (T2DM), vitamin D may influence both insulin secretion and sensitivity. An inverse relationship between T2DM and vitamin D is postulated from cross-sectional and prospective studies, though conclusive proof is as yet lacking. Available studies differ in their design and in the recommended daily allowances (RDA) of vitamin D in non-skeletal diseases and β-cell function. Large, well designed, controlled, randomized interventional studies on the potential role of vitamin D and calcium in prevention and management of T2DM are required to clarify the relationship between vitamin D and glucose homeostasis in T2DM.
INTRODUCTION
Vitamin D has long been known as the anti-rickets factor or sunshine vitamin. Today, vitamin D deficiency is considered to be a global health problem. In 2008, it was estimated that one billion individuals have vitamin D deficiency (25OH Vitamin D <20 ng/ml).1 Meanwhile, type 2 diabetes (T2DM) is one of the main non-communicable diseases. It is estimated that 366 million people had diabetes in 2011 and that by 2030 this may rise well above half a billion cases.2
Vitamin D synthesis
Vitamin D2 is ergocalciferol derived from plant and vegetable sources. Vitamin D3 is a secosteroid synthesized in the skin from 7-dehydrocholetserol (7-DHC) on exposure to sunlight by ultraviolet B-rays (UV-B rays 290-310 nm) (Figure 1),3,4 while melanin, skin pigmentation, clothing and sunscreen limit vitamin D3 production. Intensity of UV-B radiation of sunlight varies according to latitude. Hence, the farther one lives from the equator, the less UV-B is available from sunlight for induction of vitamin D synthesis in the skin.5 Vitamin D needs to be hydroxylated twice to become biologically active: it must first be converted to 25 hydroxyvitamin D3 (25OHD3) and further to 1,25(OH)2 vitamin D3 to be functionally active (Figure 1).
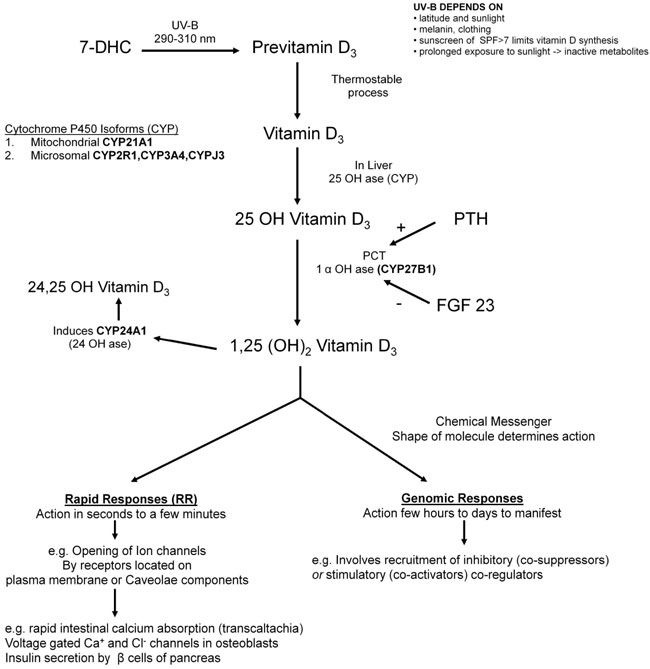
Figure 1. Vitamin D3 is secosteroid synthesized in the skin from 7-dehydrocholetserol (7-DHC) through a two step process. The B-ring of the 7-DHC is broken under UV-B rays (290-310 nm) forming pre-vitamin D3 that isomerizes to vitamin D3 in thermo-sensitive but non-enzymatic process. Vitamin D3 needs to be hydroxylated twice to become biologically active. The enzyme 25 OHase in the liver converts vitamin D3 into 25OHD.11 Serum 25OHD3 is a reliable indicator of vitamin D status.12 Vitamin D is bound to a carrier protein, the vitamin D binding protein (DBP), and transported through the circulation. In proximal tubules (PCT) of the kidney, second hydroxylation occurs by 1-α-hydroxylase (1αOHase, CYP27B1) into 1, 25 dihydroxy vitamin D3 [1,25(OH)2D3] which is the active form of vitamin D. 1,25(OH)2D3 generates biological response by the presence of its cognate receptors in selected target organs and tissues. The genomic responses of 1,25(OH)2D3 are mediated by the formation of their ligand-receptor complex with their cognate nuclear receptor. 1,25(OH)2D3 serves as a chemical messenger that transmits signals and rapid responses (RR) (e.g. opening of ion channels). A number of extra-renal sites such as immune cells epithelia of skin, gut, prostate, lung, bone, parathyroid gland and pancreatic islets may provide 1,25(OH)2D3 for local use as an intracrine or paracrine factor.13,14 Control of 1,25(OH)2D3 by non-renal tissues depends on the organ concerned. In macrophages and keratinocytes, CYP27B1 is induced by invading organisms.15
Tissue-specific actions of vitamin D
The genomic responses generally take a few hours to days to become fully manifest, the binding of 1,25(OH)2D3 ligand to VDR triggering a tight association with its heterodimeric partner retinoid X receptor (RXR). Only the 1,25(OH)2D3-RXR-VDR complex is able to penetrate the deep groove of DNA and recognize vitamin D responsive elements (VDREs) located within introns and/or at large distances from the transcription start site.6 The control of transcription requires additional recruitment of co-regulators that can be inhibitory (co-suppressors) or stimulatory (co-activators).7,8 While certain genes are selective for the co-regulator that combines with VDR and regulates their transcription, these genomic responses can be blocked by inhibitors of transcription and translation.8
1,25(OH)2D3 serves as a chemical messenger that transmits signals and rapid responses (RR) (e.g. opening of ion channels), the RR being mediated by a variety of receptors located near or associated with plasma membrane or its caveolae components.9 Caveolae are flask-shaped membrane invaginations that are enriched in spingolipids and cholesterol commonly found in both caveolae and/or lipid rafts.10 The time required for RR varies from seconds (opening of ion channels) to 10-60 minutes (e.g. activation of phosphotidylinsoitol-3’-kinases, endothelial nitric oxide synthatase). Examples of RR include rapid intestinal absorption of calcium (transcaltachia), secretion of insulin by pancreatic β-cells, opening of voltage-gated Ca+ and Cl– channels of osteoblasts and rapid migration of endothelial cells.9,10 Interestingly, one isomeric form of 1,25(OH)2D3 is used for genomic response and a different isomeric form serves as an agonist of rapid response.10 The ability of individual tissues to produce their own 1,25(OH)2D3 in a tissue-specific fashion may account for the great specificity with which vitamin D regulates numerous functions in many tissues (Figure 1).
VITAMIN D AND REGULATION OF IMMUNE FUNCTION
It is now widely recognized that the vitamin D endocrine system subserves a wide range of fundamental biological functions in cell differentiation, inhibition of cell growth as well as immunomodulation. Both forms of immunity, namely adaptive and innate, are regulated by 1,25(OH)2D3 (Figure 2). VDR are found in activated dendritic cells, macrophages and lymphocytes. These cells produce 1,25(OH)2D3 (expressing CYP27B1 which converts 25(OH)D to 1,25(OH)2D3).16 1,25(OH)2D3 regulates their proliferation and function.17,18
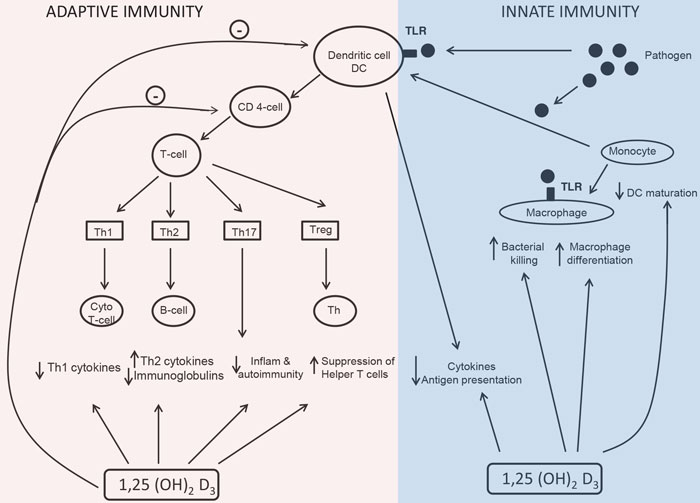
Figure 2. The action of vitamin D on macrophages is to stimulate differentiation of precursor monocyte to mature phagocytic macrophages.17,18 The dendritic cells stimulate the effector CD4+ cells to differentiate into one of the four types of T-helper cells (Th cells). Activated T-cells express VDR. 1,25(OH)2D3 inhibits the development of T-helper 1 (Th1) cells which are associated with cellular, immune response.28 1,25(OH)2D3 promotes T-helper 2 (Th2) cells associated with humoral (antibody) mediated immunity.33 Thus, 1,25(OH)2D3 promotes the T-cell shift from Th1 to Th2 cells cell function.35 1,25(OH) 2D3 inhibits the development of T helper 17 (Th17) cells which plays an essential role in combating certain pathogens and also linked to tissue damage and inflammation.35 The fourth group of CD4+ T-cells, the Treg cells exerts suppressor functions.36(Adapted from Martin Hewison, 2010 Endocrinol Metab Clin North Am 39: 365-379).
Vitamin D and innate immunity
Cytotoxic T lymphocytes (cytotoxic T-cells), Natural Killer cells (NK cells), macrophages and monocyte precursors play a central role in initial nonspecific response to tissue damage or pathogenic organisms, i.e. cell-mediated immunity. They do this by phagocytosis of cellular debris or pathogens and by assimilating the resulting waste material or eliminating it. The macrophages use the phagocytic material for antigen presentation to T-cells (Figure 2), this function involving activating toll-like receptors (TLRs) in polymorphonuclear cells (PMN).
Experimental studies have confirmed that 25(OH)D3 is able to induce intracrine VDR responses in monocytes which have been treated with TLR2/1 activator. The TLR2/1,25(OH)2D3 combination stimulates the expression of antibacterial protein cathelicidin.19 The beneficial role of vitamin D in innate immune regulations consists in its contribution to the regulation of feedback control pathways. 1,25(OH)2D3 is shown to down-regulate the expression of the monocytes TLR2 and TLR4, thereby suppressing inflammatory response by using these receptors.20 Thus, by activating CYP24 and TLR regulating mechanisms, vitamin D prevents over-elaboration of innate immune response and the damage associated with it.
Vitamin D and dendritic cells – Antigen presentation
Antigen presenting cells (APC), and specifically dendritic cells (DCs), present antigens to cells involved in the adaptive arm of the immune system. DCs, which are derived from monocytes and are heterogeneous in terms of their phenotype, function and location, are classified into two groups based on their origin: myeloid DCs (mDCs) and plasmacytoid DCs (pDCs). mDCs are the most effective antigen presenting cells, while pDCs are associated with immune tolerance. They produce different types of cytokines and chemokines and exert complimentary effects on T-cell responses.21 1,25(OH)2D3 regulates mDCs and suppresses activation of naïve T-cells.21
Dendritic cells prime CD4+ T-cells (Figure 2). DCs capture and process antigen, and once this has been effected, the DCs will mature and increase expression of co-stimulatory molecules which are necessary for interaction between the T-cell receptor and the antigen/MHC complex.22 Hampering the co-stimulatory capacity of DCs will lead to a shift from immunogenicity to tolerance. Since 1,25(OH)2D3 suppresses the expression of MHC-II molecules as well as co-stimulatory molecules,22,23 it can thus be deduced that 1,25(OH)2D3 has immunosuppressive properties. It not only inhibits maturation of DCs but also increases apoptosis of mature DCs.24 A study has shown that 1,25(OH)2D3 and its synthetic analogues inhibited the maturation of monocyte-derived DCs, thereby suppressing the capacity to present antigen to T-cells.25 Extensive proteomic analysis of DCs with 14-epivitamin D3 analogue (TX527) demonstrates that DCs are locked in an immature state and adopt a tolerogenic phenotype with a special endocytic property, as compared with mature or immature DCs.24,26 Hence, 1,25(OH)2D3 acts on dendritic cells to reduce their maturation and antigen presenting capability. Based on these data, it was proposed that vitamin D could act to promote immune tolerance, and this indeed was shown in studies of pancreatic islet cell transplantation where lower rejection rates were observed.27 This in turn was associated with enhancement of suppressor or regulatory T-cells (Tregs), the latter accounting for the fact that treatment of monocyte-derived DCs in culture with 25OHD3 suppresses DCs maturation and inhibits T-cell proliferation.27
Vitamin D and adaptive immunity
The adaptive immune response involves the ability of T-cells to produce cytokines and of B lymphocytes to produce immunoglobulins specifically combating the source of antigen presented to them by either macrophages or dendritic cells. When activated, 1,25(OH)2D3, which, exerts its inhibitory action on adaptive immunity,28 induces proliferation and differentiation of T-cells (CD4+ and CD8+) along different pathways. 1,25(OH)2D3 can induce preferential differentiation of Treg cells, a pivotal mechanism linking vitamin D and adaptive immunity29 (Figure 2). 1,25(OH)2D3 increases interlukin-10(IL-10) secretion and TLR9 expression via Treg, suggesting a link between innate and adaptive immune response.30 The effect of vitamin D on CD8+ suppressor T-cells is limited. However, 1,25(OH)2D3 actively regulates cytokine production by CD8+ cells and regulates proliferation of CD8+ cells following specific antigen stimuli.31
1,25(OH)2D3 inhibits cytokine secretion associated with Th1(IFN-γ) cells.28 It also inhibits IL-12a T-cell stimulatory factor involved in differentiation of naïve T-cells into Th0 cells which further develop into Th1 cell or Th2 cells.32 Vitamin D down-regulates pro-inflammatory cytokines, such as IL-2, IL-6, IFN-γ and tumor necrosis factor (TNF)-β while enhancing anti-inflammatory cytokines like IL-4, IL-10 and TGF-β.33
Active B lymphocytes express VDR, while resting B lymphocytes do not. 1,25(OH)2D3 decreases proliferation of B lymphocytes and immunoglobulin and induces apotosis.35 Given that indirect mediation by T-cells and monocytes or macrophages are the most important mechanisms of action,34,17 hormonal vitamin D retards differentiation of β-cell precursors into plasma cells and suppresses proliferation and immunoglobulin production. This is beneficial for conditions in which the immune system is directed against the body’s own tissues, i.e. autoimmunity (Figure 2).
DIABETES MELLITUS AND VITAMIN D SUPPLEMENTATION
A PubMed search was conducted using the terms “Vitamin D-insulin resistance–type 1 and 2 diabetes mellitus”, “Vitamin D insulin–sensitivity–type 1 and 2 diabetes mellitus”, “Vitamin D insulin–secretion–type 1 and 2 diabetes mellitus” under the category of clinical trials from the year 1985 till now. These references are tabulated in the present review.
Vitamin D in type 1 Diabetes Mellitus
Type 1 diabetes mellitus (T1DM) is a chronic disease of multifactorial nature resulting from progressive autoimmune destruction of pancreatic islet cells at the early stages of disease.37 Genetic predisposition and environmental factors are both contributory to the development of the disease. It is hypothesized that vitamin D may have a therapeutic role in T1DM via its immune-modulatory properties. More specifically, at the time of the onset of T1DM, chronic inflammatory insulinitis (infiltrates) is found in the islets of Langerhans involving CD8+ and CD4+ T-cells, B lymphocytes and macrophages.37 Vitamin D inhibits the production of IFN-γ and IL-2 cytokines, known to activate both macrophages and cytotoxic T-cells, which leads to destruction of pancreatic islet cells.38 Furthermore, vitamin D suppresses APC and modulates development of CD4+ lymphocytes.39 Exposure of pancreatic β-cells to proinflammatory cytokine induces endoplasmic reticulum stress and apotosis.40 1,25(OH)2D3 protects β-cells by reducing exposure of MHC-1 molecules.41 It induces antiapoptotic A20 protein and decreases exposure of a transmembrane cell surface receptor, thus transducing apoptotic death signals, thereby contributing to the pathogenesis of T1DM, namely, Fas.42,43 Ecologically, observational studies have been conducted that showed a north-south gradient and that have linked the incidence of T1DM with latitude and seasonal pattern,44 suggesting an inverse correlation between effective exposure of sunlight and incidence of T1DM.
Animal experiments
As demonstrated in animal models of T1DM non-obese diabetic mice (NOD mice), high doses of 1,25(OH)2D3(5 µg/kg/alternate day) suppresses insulitis and diabetes development, this accompanied by a diminished number of effector T-cells as well as induction of Treg cells, which was shown to be the basis of protection.43,45 In an inflammation-driven model of diabetes (streptozotocin-induced diabetes model), 1,25(OH)2D3 reduced the incidence of diabetes but had little effect in reverting overt diabetes.46 Pharmacological doses of 1,25(OH)2D3 lead to hypercalcemia and bone calcification.46 Use of structural analogues of 1,25(OH)2D3 could partly solve this issue. It was recently observed that a combination of 14-epivitamin D3 analogue (TX527) with cyclosporine (IFN-β) induced delay in recurrence of diabetes after islet transplantation with increased expression of IL-10 in islet cell grafts.46 An analogue of vitamin D (KH1060) has been shown to prevent the onset of diabetes in the NOD mouse. Overall, animal studies suggest a halt of the progression of T1DM by vitamin D analogues even after initiation of an autoimmune attack, since administration of MC 1288 with or without cyclosporine A in established insulitis resulted in reduction of diabetes incidence.47
Studies in humans
Small-scale epidemiological clinical interventions have produced interesting findings. High doses of vitamin D (2000 IU/day) during the first year of life reduced the risk of development of T1DM.48 Studies by Stein et al48 have demonstrated that use of cod liver oil from 7-12 months of life was associated with lowered risk of developing T1DM in later life.48 Also, cod liver oil taken by pregnant mothers in the 3rd trimester of pregnancy was associated with decreased risk of T1DM.48 The European Community-sponsored Concerted Action on the Epidemiology and Prevention of Diabetes (EURODIAB) revealed that there was a 33% risk reduction (OR 0.67) of T1DM in children who received vitamin D supplementation in earlier life compared to non-supplemented subjects.49,50 A meta-analysis of four case-control studies and one cohort study showed a 29% risk reduction in infants who were supplemented with vitamin D as compared with those who were not [pooled odds ratio(OR) 0.71, 95% confidence intervals (CI) 0.60-0.84].49,50 The screening for T1DM genetic risk in newborns of an Italian study (Prevefin Italy)51 sought to validate the efficacy of two primary prevention strategies (vitamin D supplementation and β-casein-free diet) in preventing autoimmune aggression of pancreatic β-cells and ultimately the onset of T1DM. Cord blood samples of 9409 Caucasian newborns were screened for risk of T1DM and HLA class II markers. Seventy-three newborns qualified for the study. Informed consent was obtained for those with a high genetic risk of T1DM and, after β-cell autoantibody analysis (GAD antibodies, IAA, IA-2A), were randomized into two arms: one arm to receive 500 IU of vitamin D free diet and the second arm to receive 500 IU of vitamin D plus β-casein-free diet in the first 12 months of life. Every three months, apart from clinical and auxiological examination, antibody analysis was also carried out. If antibodies were positive, they were metabolically tested with the glucagon test. The preliminary study showed a low rate of transient or persistent antibody positivity. Long-term results are awaited.51 Studies in patients with Latent Autoimmune Diabetes in Adults (LADA) developing T1DM during treatment with analogues of vitamin D in addition to insulin treatment exhibited a better ability to preserve β-cell function compared with those treated with insulin alone.52 Studies in pregnant women have displayed these beneficial effects in the form of reduced occurrence of diabetes-specific autoantibodies in offspring of mothers supplemented with vitamin D.53
While the immune protective effect of vitamin D is thought to be mainly based on Treg cells, these therefore constituting a central player in maintenance of self-tolerance, there are an insufficient number of studies on the effect of vitamin D supplementation on peripheral T-cells. Recent studies of Bock et al54 showed that supplementation with oral vitamin D at doses of 140,000 IU monthly for three months significantly increased the Tregs after this short period. These results indicate that vitamin D may be a useful therapeutic agent exerting immune modulatory effects that involve a stimulatory effect on Tregs. Table 1 presents randomized clinical trials that studied the effect of vitamin D on established T1DM and their outcomes.55-59
In summary, the proven immune modulatory effect of vitamin D strongly points to a rational hypothesis for its role in the pathogenesis of type 1 diabetes, one of the most common autoimmune diseases, this being is adequately backed by animal studies and ample epidemiological and observational data in humans. However, no large prospective randomized control studies are available to support a therapeutic role of vitamin D agent in preventing or postponing the disease (Table 1). Hence for now, pragmatism dictates ensuring adequate levels and avoiding deficiency of vitamin D in vulnerable groups as the key priority in day-to-day clinical practice.
Vitamin D in type 2 Diabetes Mellitus
Mechanisms
The main defects that determine the development of T2DM are insulin resistance, pancreatic β-cell dysfunction and systemic inflammation.
β-cell function and insulin secretion
Vitamin D promotes pancreatic β-cell function in numerous ways.
1. Direct actions: Activation of vitamin D occurs in pancreatic β-cells by intracellular 1-α-OHase enzyme. Vitamin D enhances insulin secretion and promotes β-cell survival by modulating the generation and effects of cytokines. The anti-apoptotic action of vitamin D is mediated by down-regulating Fas-related pathways (Fas/Fas-L) (Figure 3).60,61
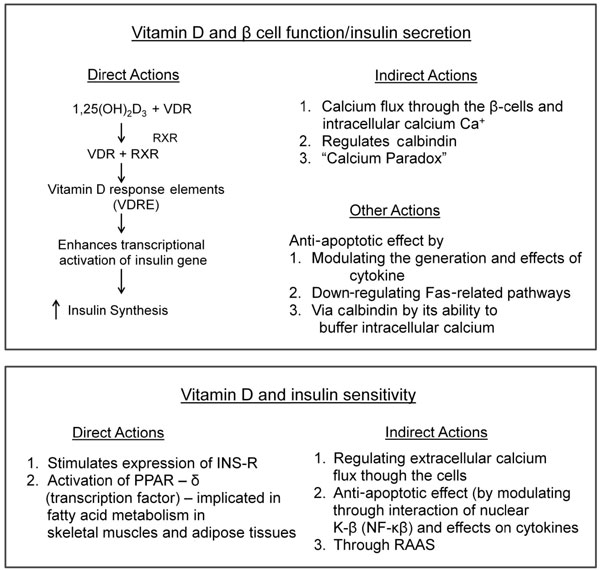
Figure 3. 1,25(OH)2D3 enhances insulin secretion by interacting with the 1,25(OH)2D3-RXR-VDR complex which binds to vitamin D responsive elements (VDRE) found in the insulin gene promoter region, to enhance the transcriptional activation of the insulin gene and increase insulin synthesis.64 It acts as a modulator of depolarization – stimulated insulin release via regulation of intracellular calcium. INS-R: insulin receptor.
2. Indirect actions: Insulin secretion is a calcium-dependent process and is influenced by calcium flux through the cell membrane by RR.60 Vitamin D regulates calbindin, a cytosolic calcium-binding protein found in β-cells. It acts as a modulator of depolarization-stimulated insulin release via regulation of intracellular calcium (Figure 3). Thus, vitamin D could indirectly effect insulin secretion additionally by regulating calbindin. Another plausible mechanism could be one whereby low vitamin D status induces secondary hyperparathyroidism (SHPT). The raised parathyroid hormone (PTH) inhibits insulin synthesis and secretion in β-cells and insulin resistance in target cells by regulating intracellular calcium (Figure 3). The SHPT may actually cause a paradoxical increase in intracellular calcium and in turn may impair the calcium signal needed for glucose induced insulin secretion, this is known as the “calcium paradox”.62
Vitamin D and insulin sensitivity
In insulin responsive tissues, such as skeletal muscle and adipose tissue, calcium is essential for insulin-mediated intracellular processes. Indeed, a narrow range of intracellular calcium is needed for optimal function.63 Vitamin D enhances insulin sensitivity by stimulating the expression of insulin receptors and/or by activating peroxisome proliferator-activated receptor-δ (PPAR- δ).64 PPAR-δ is implicated in the regulation of fatty acid metabolism in skeletal muscles and adipose tissue (Figure 3). The indirect effect of vitamin D is exerted by regulating calcium flux through the cell membrane and intracellular calcium by RR (Figures 1 and 3). Changes in intracellular calcium in insulin target tissues may contribute to peripheral insulin resistance via impaired signal transduction pathways leading to decreased glucose transporter activity. Vitamin D may promote β-cells survival by inactivation of nuclear factor-κB (NF-κb) and effects of cytokines.64 Vitamin D could also affect insulin resistance indirectly through the rennin-angiotensin-aldosterone system (RAAS). Angiotensin II inhibits the action of insulin in vascular and skeletal muscle tissue leading to impaired glucose uptake. Vitamin D suppresses rennin formation and local pancreatic RAAS. Hence, vitamin D could be a negative endocrine regulator of RAAS.65
Vitamin D and systemic inflammation
According to the current understanding of the pathogenesis of T2DM, inflammation is postulated to play an important role in insulin resistance, while β-cell function may be affected via cytokine-induced apoptosis. Vitamin D is thought to modulate the expression and activity of cytokines and protect the β-cell against cytokine-induced apoptosis, one such effect being the countering of cytokine-induced Fas expression.60,61 Other immune modulating effects of vitamin D include blockade of dendritic cell differentiation, inhibition of lymphocyte proliferation, enhanced regulation of T-lymphocytes, development and down-regulation of cytokine expression. These immune modulatory effects of vitamin D might provide additional protection against inflammation-triggered worsening of insulin resistance and, potentially, β-cell function.
Animal studies
Studies in ob/ob mouse showed that 1,25(OH)2D3 treatment improved hyperglycemia, hyperinsulinemia and fat tissue responsiveness to hormones.69 Vitamin D supplementation had beneficial effects in obese spontaneously hypertensive rats (SHR) and Wistar rats, where there was a reduction in glucose levels in vitamin D supplemented animals.66 Glucose-mediated insulin secretion from β-cells was impaired in vitamin D deficient rats.67 Insulin secretion was restored on vitamin D supplementation.67-69
Studies in humans
Seasonal variations of glycemic control in patients with T2DM, with a worsening in winter, was explained partly by fluctuations in 25OHD3 concentrations on exposure to UV radiation.44
1. Cross-sectional studies
Several cross-sectional studies have examined the association between serum 25(OH)D3 concentrations and prevalence of T2DM. Most studies have reported an inverse association between 25(OH)D3 levels and T2DM,70 while others failed to show such associations (Table 2).71 Large population-based studies such as the Third National Health and Nutrition Examination Survey (NHANES III) have disclosed a positive relationship between serum 25(OH)D3 levels and insulin sensitivity.72 In this study, serum 25(OH)D3 levels were inversely associated with metabolic syndrome in both genders and all three major racial and ethnic groups (non-Hispanic whites, Mexican Americans and the remaining groups of American adults). Further studies using NHANES III data has confirmed inverse associations between serum 25(OH)D3 concentrations and fasting hyperglycemia as well as insulin resistance.71,72 In a birth cohort study, serum 25(OH)D3 concentrations were inversely associated with elevated glycosylated hemoglobin levels.73
These observational studies are also supported by laboratory studies. In one study,74 there was a positive correlation between insulin sensitivity, as measured by hyperglycemic clamp, and 25(OH)D3 concentrations even after correcting for confounding factors such as body mass index (r=0.25; P=0.007). There are, however, a number of limitations of many cross-sectional studies. To mention a few, studies investigating the relationship between 25(OH)D3 levels and insulin secretion sensitivity have used indirect primary measures, such as fasting insulin and HOMA-R, QUICKI,75 even though the use of hyperglycemic clamp studies is the gold standard for testing these parameters. In a recent study in postmenopausal women (n=753), 25(OH)D3 concentrations were inversely correlated with fasting plasma glucose (r= -0.15; P=0.001).76
2. Prospective studies
There are relatively few prospective studies on the association between T2DM and vitamin D status. In the Women’s Health Study, a vitamin D intake of 511 IU/day or more was associated with less incident T2DM compared to that observed on 159 IU/day or less.77 In one of the largest prospective studies, the Nurses’ Health Study (USA), women with the highest vitamin D intake, i.e. greater than 800 IU/day, had 33% lower risk of incident diabetes compared to women with intakes of less than 200 IU/day.78 In the Mini-Finland Health Study, the relative risk of T2DM was 0.6 in subjects with the highest quintiles of 25(OH)D3 (70.9 nmol/l) compared to those with the lowest quintiles (22.4 nmol/l; P<0.01). Pooled data from two nested case-control studies by the Finnish Mobile Clinic showed that the men with the highest serum 25(OH)D3 quintiles (69.11 nmol/l) vs those with the lowest quintiles (22.3 nmol/l; P<0.001) had 82% reduced risk of T2DM incidence.89
Role of calcium intake along with vitamin D in T2DM
In the Women’s Health Study, calcium intake (after adjustment of vitamin D intake) was inversely associated with prevention of metabolic syndrome.80 Women with the highest calcium (>1200 mg/day) and vitamin D (>800 IU/day) intake had 33% lower risk of T2DM compared to women with the lowest calcium (<600 mg/day) and vitamin D (<400 IU/day) intake daily.77
3. Intervention studies
There are several interventional studies (Table 2) on the effect of vitamin D supplementation on T2DM and the effect of combined vitamin D and calcium supplementation on T2DM. It is difficult to draw definitive conclusions from these trials. There are reports of the improvement of insulin secretion six months after supplementation of vitamin D in vitamin D deficient T2DM subjects.70 Other studies have reported significant improvement in insulin secretion after variable doses and length of vitamin D supplementation in subjects with T2DM.66 Generally speaking, the available studies are limited by a lack of randomized, placebo-controlled dosages and non-reporting of serum 25(OH)D to attain sufficient vitamin D concentrations.
In summary, direct actions of vitamin D promoting β-cell survival and its action on skeletal and adipose tissue that are shown to improve insulin sensitivity are compelling reasons to consider its potential in positively influencing glycemic status in patients with type 2 diabetes. Although an abundance of observational data from the Nurses’ Health Study supported the hypothesis of lesser prevalence of hyperglycemia with adequate intake of calcium and vitamin D, equally significant data from a large cohort involving postmenopausal women in the Women’s Health Initiative Study in the USA, conducted over a period of 7 years, discredited this positive association. Numerous studies across the globe involving small numbers of patients for shorter durations have yielded similarly conflicting conclusions (Table 2). A recent study has shown that optimizing serum 25(OH)D concentrations and supplementation of calcium in vitamin D deficient individuals improves fasting plasma glucose and pancreatic β-cell reserve (by HOMA model).109
Whilst awaiting well-designed, long-term, prospective, randomized intervention trials involving large groups of patients across the globe which will finally resolve the several conflicting issues, the most prudent practice to be undertaken in everyday clinical practice is to ensure adequate vitamin D levels and dietary calcium intake.
CONCLUSIONS
While bearing in mind that vitamin D regulates both innate and adaptive immunity, thus indicating its potential role in preventing and treating T1DM, an autoimmune disorder of pancreatic islets leading to progressive destruction of insulin secreting β-cells, it is also evident that vitamin D deficiency cannot be the sole determining factor of T1DM. Clearly, both genetic and environmental factors determine the development of the disease. From the existing data, it could be postulated that optimal doses of vitamin D supplementation may influence the pattern of immune regulation and subsequent progression to T1DM in a genetically susceptible individual. However, more studies are required to substantiate the relation between T1DM and vitamin D/vitamin D analogues in the pattern of immune regulations in susceptible individuals.
Multiple studies strongly suggest a role of vitamin D in the wellbeing of β-cells, insulin production and secretion, tissue sensitivity to insulin and the susceptibility to T2DM. An inverse relationship between T2DM and vitamin D is suggested by cross-sectional and prospective studies pointing to a direct link between the risk of T2DM and vitamin D. However, conclusive proof is as yet lacking, this mainly owing to the fact most of the study results have been limited by study design. Thus, large, well designed, controlled, randomized interventional studies of vitamin D and calcium in potential prevention and management of T2DM are urgently required to clarify the relationship between vitamin D and glucose homeostasis in T2DM.
REFERENCES
1. Chagas CE, Borges MC, Martini LA, Rogero MM, 2012 Focus on vitamin D, inflammation and type 2 diabetes. Nutrients 4: 52-67.
2. IDF diabetes atlas. International Diabetes Federation 2009 Available at: http://www.idf.org/diabetesatlas/5e/the-global-burden last accessed on 30/09/2012.
3. Reichrath J, 2007 Exp Dermatol 16: 618-625.
4. Holick MF, 2004 Vitamin D: importance in the prevention of cancers, type 1 diabetes, heart disease, and osteoporosis. Am J Clin Nutr 79: 362-371.
5. Webb AR, Kline L, Holick MF, 1988 Influence of season and latitude on the cutaneous synthesis of vitamin D3: exposure to winter sunlight in Boston and Edmonton will not promote vitamin D3 synthesis in human skin. J Clin Endocrinol Metab 67: 373-378.
6. Bikle DD, Pillai S, Gee E, Hincenbergs M, 1985 Regulation of 1,25-dihydroxyvitamin D production in human keratinocytes by interferon-gamma. Endocrinology 124: 655-660.
7. Ohyama Y, Ozono K, Uchida M, et al, 1996 Functional assessment of two vitamin D-responsive elements in the rat 25- hydroxyvitamin D3 24-hydroxylase gene. J Biol Chem 271: 30381-30385.
8. DeLuca HF, 2004 Overview of general physiologic features and functions of vitamin D. Am J Clin Nutr 80: Suppl 6: 1689-1696.
9. McKenna NJ, Lanz RB, O’Malley BW, 1999 Nuclear receptor coregulators: cellular and molecular biology. Endocr Rev 20: 321-344.
10. Razani B, Woodman SE, Lisanti MP, 2002 Caveolae: from cell biology to animal physiology. Pharmacol Rev 54: 431-467.
11. Prosser DE, Jones G, 2004 Enzymes involved in the activation and inactivation of vitamin D. Trends Biochem Sci 29: 664-673.
12. Takiishi T, Gysemans C, Bouillon R, Mathieu C, 2010 Vitamin D and diabetes. Endocrinol Metab Clin North Am 39: 419-446.
13. Bikle DD, 2009 Extra renal synthesis of 1,25 dihydroxy vitamin D and its health implications. Clin Rev Bone Miner Metab 7: 114-125.
14. Nishimura A, Shinki T, Jin CH, et al, 1994 Regulation of messenger ribonucleic acid expression of 1 alpha,25- dihydroxyvitamin D3-24-hydroxylase in rat osteoblasts. Endocrinology 134: 794-1799.
15. Schauber J, Dorschner RA, Coda AB, et al, 2007 Injury enhances TLR2 function and antimicrobial peptide expression through a vitamin D-dependent mechanism. J Clin Invest 117: 803-811.
16. Nagpal S, Na S, Rathnachalam R, 2005 Noncalcemic actions of vitamin D receptor ligands. Endocr Rev 26: 662-687.
17. Chen S, Sims GP, Chen XX, Gu YY, Chen S, Lipsky PE, 2007 Modulatory effects of 1,25-dihydroxyvitamin D3 on human B cell differentiation. J Immunol 179: 1634-1647.
18. Bikle DD, 2009 Vitamin D and immune function: Understanding common pathways. Curr Osteoporos Rep 7: 58-63.
19. Tanaka H, Abe E, Miyaura C, Shiina Y, Suda T, 1983 1-alpha,25-dihydroxyvitamin D3 induces differentiation of human promyelocytic leukemia cells (HL-60) into monocyte-macrophages, but not into granulocytes. Biochem Biophys Res Commun 117: 86-92.
20. Liu PT, Stenger S, Li H, et al, 2006 Toll-like receptor triggering of a vitamin D-mediated human antimicrobial response. Science 311: 1770-1773.
21. Liu YJ, 2005 IPC: professional type 1 interferon-producing cells and plasmacytoid dendritic cell precursors. Annu Rev Immunol 23: 275-306.
22. Guery JC, Adorini L, 1995 Dendritic cells are the most efficient in presenting endogenous naturally processed self-epitopes to class II-restricted T cells. J Immunol 154: 536-544.
23. Schwartz RH, 1990 A cell culture model for T lymphocyte clonal anergy. Science 248: 1349-1356.
24. Penna G, Adorini L, 2000 1-Alpha,25-dihydroxyvitamin D3 inhibits differentiation, maturation, activation, and survival of dendritic cells leading to impaired alloreactive T cell activation. J Immunol 164: 2405-2411.
25. Sadeghi K, Wessner B, Laggner U, et al, 2006 Vitamin D3 down-regulates monocyte TLR expression and triggers hyporesponsiveness to pathogen-associated molecular patterns. Eur J Immunol 36: 361-371.
26. van Halteren AG, Tysma OM, van Etten E, Mathieu C, Roep BO, 2004 1-alpha,25-dihydroxyvitamin D3 or analogue treated dendritic cells modulate human autoreactive T cells via the selective induction of apoptosis. J Autoimmun 23: 233-239.
27. Griffin MD, Lutz WH, Phan VA, et al, 2000 Potent inhibition of dendritic cell differentiation and maturation by vitamin D analogs. Biochem Biophys Res Commun 270: 701-708.
28. Lemire JM, Adams JS, Sakai R, Jordan SC, 1984 1-alpha,25-dihydroxyvitamin D3 suppresses proliferation and immunoglobulin production by normal human peripheral blood mononuclear cells. J Clin Invest 74: 657-661.
29. Mathieu C, Badenhoop K, 2005 Vitamin D and type 1 diabetes mellitus: state of the art. Trends Endocrinol Metab 16: 261-266.
30. Spach KM, Nashold FE, Dittel BN, Hayes CE, 2006 IL-10 signaling is essential for 1,25-dihydroxyvitamin D3- mediated inhibition of experimental autoimmune encephalomyelitis. J Immunol 177: 6030-6037.
31. Iho S, Iwamoto K, Kura F, Okuno Y, Takahashi T, Hoshino T, 1990 Mechanism in 1,25(OH)2D3-induced suppression of helper/suppressor function of CD4/CD8 cells to immunoglobulin production in B cells. Cell Immunol 127: 12-25.
32. Boonstra A, Barrat FJ, Crain C, Heath VL, Savelkoul HF, O’Garra A, 2001 1alpha,25-Dihydroxyvitamin D3 has a direct effect on naive CD4(1) T cells to enhance the development of Th2 cells. J Immunol 167: 4974-4980.
33. Imazeki I, Matsuzaki J, Tsuji K, Nishimura T, 2006 Immunomodulating effect of vitamin D3 derivatives on type-1 cellular immunity. Biomed Res 27: 1-9.
34. van Etten E, Mathieu C, 2005 Immunoregulation by 1,25-dihydroxyvitamin D3: basic concepts. J Steroid Biochem Mol Biol 97: 93-101.
35. Overbergh L, Decallonne B, Waer M, et al, 2009 1alpha,25-dihydroxyvitamin D3 induces an autoantigen specific T-helper 1/T-helper 2 immune shift in NOD mice immunized with GAD65 (p524-543). Diabetes 49: 1301-1307.
36. Korn T, Oukka M, Kuchroo V, Bettelli E, 2007 Th17 cells: effector T cells with inflammatory properties. Semin Immunol 19: 362-371.
37. Atkinson MA, Maclaren NK, 1995 The pathogenesis of insulin-dependent diabetes mellitus. Engl J Med 331: 1428-1436.
38. Foulis AK, McGill M, Farquharson MA, 1991 Insulitis in type 1 (insulin-dependent) diabetes mellitus in man–macrophages, lymphocytes, and interferon-gamma containing cells. J Pathol 165: 97-103.
39. Riachy R, Vandewalle B, Belaich S, et al, 2001 Beneficial effect of 1,25 dihydroxyvitamin D3 on cytokine-treated human pancreatic islets. J Endocrinol 169: 161-168.
40. Mauricio D, Mandrup-Poulsen T, Nerup J, 1996 Vitamin D analogues in insulin-dependent diabetes mellitus and other autoimmune diseases: a therapeutic perspective. Diabetes Metab Rev 12: 57-68.
41. Eizirik DL, Cardozo AK, Cnop M, 2008 The role for endoplasmic reticulum stress in diabetes mellitus. Endocr Rev 29: 42-61.
42. Hahn HJ, Kuttler B, Mathieu C, Bouillon R, 1997 1,25-Dihydroxyvitamin D3 reduces MHC antigen expression on pancreatic beta-cells in vitro. Transplant Proc 29: 2156-2157.
43. Riachy R, Vandewalle B, Moerman E, et al, 2006 1,25-Dihydroxyvitamin D3 protects human pancreatic islets against cytokine-induced apoptosis via down-regulation of the Fas receptor. Apoptosis 11:151-159.
44. Mohr SB, Garland CF, Gorham ED, Garland FC, 2008 The association between ultraviolet B irradiance, vitamin D status and incidence rates of type 1 diabetes in 51 regions worldwide. Diabetologia 51: 1391-1398.
45. Casteels K, Waer M, Laureys J, et al, 1998 Prevention of autoimmune destruction of syngeneic islet grafts in spontaneously diabetic nonobese diabetic mice by a combination of a vitamin D3 analog and cyclosporine. Diabetes 51: 1367-1374.
46. Del Pino-Montes J, Benito GE, Fernandez-Salazar MP, et al, 2004 Calcitriol improves streptozotocin-induced diabetes and recovers bone mineral density in diabetic rats. Calcif Tissue Int 75: 526-532.
47. Casteels K, Waer M, Laureys J, et al, 1998 Prevention of autoimmune destruction of syngeneic islet grafts in spontaneously diabetic nonobese diabetic mice by a combination of a vitamin D3 analog and cyclosporine. Transplantation 65: 1225-1232.
48. Zipitis CS, Akobeng AK, 2008 Vitamin D supplementation in early childhood and risk of type 1 diabetes: a systematic review and meta-analysis. Arch Dis Child 93: 512-517.
49. Vitamin D supplementation in early childhood and risk for Type I (insulin-dependent) diabetes mellitus. The EURODIAB Substudy 2 Study Group 1999 Diabetologia 42: 51-54.
50. EURODIAB ACE Study Group, 2000 Variation and trends in incidence of childhood diabetes in Europe. Lancet 355: 873-876.
51. Lorini R, Minicucci L, Napoli F, et al, 2005 Screening for type 1 diabetes genetic risk in newborns of continental Italy. Primary prevention (Prevefin Italy)–preliminary data. Acta Biomed 76(Suppl 3): 31-35.
52. Li X, Liao L, Yan X, et al, 2009 Protective effects of 1-alpha-hydroxyvitamin D3 on residual beta-cell function in patients with adult-onset latent autoimmune diabetes. (LADA). Diabetes Metab Res Rev 25: 411-416.
53. Brekke HK, Ludvigsson J, 2007 Vitamin D supplementation and diabetes-related autoimmunity in the ABIS study. Pediatr Diabetes 8: 11-14.
54. Bock G, Prietl B, Mader JK, et al, 2011 The effect of vitamin D supplementation on peripheral regulatory T cells and β cell function in healthy humans: a randomized controlled trial. Diabetes Metab Res Rev 27: 942-945.
55. Gabbay MA, Sato MN, Finazzo C, Duarte AJ, Dib SA, 2012 Effect of cholecalciferol as adjunctive therapy with insulin on protective immunologic profile and decline of residual β-cell function in new-onset type 1 diabetes mellitus. Arch Pediatr Adolesc Med 166: 601-607.
56. Aljabri KS, Bokhari SA, Khan MJ, 2010 Glycemic changes after vitamin D supplementation in patients with type 1 diabetes mellitus and vitamin D deficiency. Ann Saudi Med 30: 454-458.
57. Bizzarri C, Pitocco D, Napoli N, IMDIAB Group, et al, 2010 No protective effect of calcitriol on beta-cell function in recent-onset type 1 diabetes: the IMDIAB XIII trial. Diabetes Care 33: 1962-1963.
58. Walter M, Kaupper T, Adler K, Foersch J, Bonifacio E, Ziegler AG, 2010 No effect of the 1alpha,25-dihydroxyvitamin D3 on beta-cell residual function and insulin requirement in adults with new-onset type 1 diabetes. Diabetes Care 33: 1443-1448.
59. Pitocco D, Crinò A, Di Stasio E, IMDIAB Group, et al, 2006 The effects of calcitriol and nicotinamide on residual pancreatic beta-cell function in patients with recent-onset Type 1 diabetes (IMDIAB XI). Diabet Med 23: 920-923.
60. Norman AW, 2006 Minireview: vitamin D receptor: new assignments for an already busy receptor. Endocrinology 147: 5542-5548.
61. Eliades M, Pittas AG, 2010 Vitamin D and type 2 diabetes. In Vitamin D Physiology, Molecular Biology and clinical applications ed. Holick M F. Humana press: pp, 895-920.
62. Fujita T, Palmieri GM, 2000 Calcium paradox disease: calcium deficiency prompting secondary hyperparathyroidism and cellular calcium overload. J Bone Miner Metab 18: 109-125.
63. Fujita T, 2000 Calcium paradox: consequences of calcium deficiency manifested by a wide variety of diseases. J Bone Miner Metab 18: 234-236.
64. Dunlop TW, Väisänen S, Frank C, Molnár F, Sinkkonen L, Carlberg C, 2005 The human peroxisome proliferator-activated receptor delta gene is a primary target of 1alpha,25-dihydroxyvitamin D3 and its nuclear receptor. J Mol Biol 349: 248-260.
65. Wei Y, Sowers JR, Clark S.E, Li W, Ferrario CM, Stump CS, 2008 Angiotensin II- induced skeletal muscle insulin resistance mediated by NF-kappaB activation via NADPH oxidase. Am J Physiol Endocrinol Metab 294: E345-351.
66. Gregori S, Giarratana N, Smiroldo S, Uskokovic M, Adorini L, 2002 1alpha,25-dihydroxyvitamin D3 analog enhances regulatory T-cells and arrests autoimmune diabetes in NOD mice. Diabetes 51: 1367-1374.
67. Chertow BS, Sivitz WI, Baranetsky NG, Cordle MB, DeLuca HF, 1986 Islet insulin release and net calcium retention in vitro in vitamin D-deficient rats. Diabetes 35: 771-775.
68. Cade C, Norman AW, 1987 Rapid normalization/stimulation by 1,25-dihydroxyvitamin D3 of insulin secretion and glucose tolerance in the vitamin D–deficient rat. Endocrinology 120: 1490-1497.
69. Norman AW, Frankel JB, Heldt AM, Grodsky GM, 1980 Vitamin D deficiency inhibits pancreatic secretion of insulin. Science 209: 823-825.
70. Pittas AG, Lau J, Hu FB, Dawson-Hughes B, 2007 The role of vitamin D and calcium in type 2 diabetes. A systematic review and meta-analysis. J Clin Endocrinol Metab 92: 2017-2029.
71. Martins D, Wolf M, Pan D, et al, 2007 Prevalence of cardiovascular risk factors and the serum levels of 25-hydroxyvitamin D in the United States: data from the Third National Health and Nutrition Examination Survey. Arch Intern Med 167: 1159-1165.
72. Scragg R, Sowers M, Bell C, 2004 Third National Health and Nutrition Examination Survey. Serum 25-hydroxyvitamin D, diabetes, and ethnicity in the Third National Health and Nutrition Examination Survey. Diabetes Care 27: 2813-2818.
73. Hyppönen E, Boucher BJ, Berry DJ, Power C, 2008 25-hydroxyvitamin D, IGF-1, and metabolic syndrome at 45 years of age: a cross-sectional study in the 1958 British Birth Cohort. Diabetes 57: 298-305.
74. Chiu KC, Chu A, Go VL, Saad MF, 2004 Hypovitaminosis D is associated with insulin resistance and beta cell dysfunction. Am J Clin Nutr 79: 820-825.
75. Alvarez J.A, Ashraf A, 2010 Role of vitamin D in insulin secretion and insulin sensitivity for glucose homeostasis. Int J Endocrinol 2010: 351-385.
76. Need AG, O’Loughlin PD, Horowitz M, Nordin BE, 2005 Relationship between fasting serum glucose, age, body mass index and serum 25 hydroxyvitamin D in postmenopausal women. Clin Endocrinol (Oxf) 62: 738-741.
77. Liu S, Song Y, Ford ES, Manson JE, Buring JE, Ridker PM, 2005 Dietary calcium, vitamin D, and the prevalence of metabolic syndrome in middle-aged and older U.S. women. Diabetes Care 28: 2926-2932.
78. Pittas AG, Dawson-Hughes B, Li T, et al, 2006 Vitamin D and calcium intake in relation to type 2 diabetes in women. Diabetes Care 29: 650-656.
79. Knekt P, Laaksonen M, Mattila C, et al, 2008 Serum vitamin D and subsequent occurrence of type 2 diabetes. Epidemiology 19: 666-671.
80. Harris SS, Pittas AG, Palermo NJ, 2012 A randomized, placebo-controlled trial of vitamin D supplementation to improve glycaemia in overweight and obese African Americans. Diabetes Obes Metab 14: 789-794.
81. Eftekhari MH, Akbarzadeh M, Dabbaghmanesh MH, et al, 2011 Impact of treatment with oral calcitriol on glucose indices in type 2 diabetes mellitus patients. Asia Pac J Clin Nutr 20: 521-526.
82. Mitri J, Dawson-Hughes B, Hu FB, Pittas AG, 2011 Effects of vitamin D and calcium supplementation on pancreatic β cell function, insulin sensitivity, and glycemia in adults at high risk of diabetes: the Calcium and Vitamin D for Diabetes Mellitus (CaDDM) randomized controlled trial. Am J Clin Nutr 94: 486-494.
83. Nikooyeh B, Neyestani TR, Farvid M, et al, 2011 Daily consumption of vitamin D- or vitamin D + calcium-fortified yogurt drink improved glycemic control in patients with type 2 diabetes: a randomized clinical trial. Am J Clin Nutr 93: 764-771.
84. Sabherwal S, Bravis V, Devendra D, 2010 Effect of oral vitamin D and calcium replacement on glycaemic control in South Asian patients with type 2 diabetes. Int J Clin Pract 64: 1084-1089.
85. von Hurst PR, Stonehouse W, Coad J, 2010 Vitamin D supplementation reduces insulin resistance in South Asian women living in New Zealand who are insulin resistant and vitamin D deficient – a randomized, placebo-controlled trial. Br J Nutr 103: 549-555.
86. Pittas AG, Harris SS, Stark PC, Dawson-Hughes B, 2007 The effects of calcium and vitamin D supplementation on blood glucose and markers of inflammation in nondiabetic adults. Diabetes Care 30: 980-986.
87. Borissova AM, Tankova T, Kirilov G, Dakovska L, Kovacheva R, 2003 The effect of vitamin D3 on insulin secretion and peripheral insulin sensitivity in type 2 diabetic patients. Int J Clin Pract 57: 258-261.
88. Boucher BJ, Mannan N, Noonan K, Hales CN, Evans SJ, 1995 Glucose intolerance and impairment of insulin secretion in relation to vitamin D deficiency in East London Asians. Diabetologia 38: 1239-1245.
89. Orwoll E, Riddle M, Prince M, 1994 Effects of vitamin D on insulin and glucagon secretion in non-insulin-dependent diabetes mellitus. Am J Clin Nutr 59: 1083-1087.
90. Inomata S, Kadowaki S, Yamatani T, Fukase M, Fujita T, 1986 Effect of 1α(OH)- vitaminD3 on insulin secretion in diabetes mellitus. Bone Miner 1: 187-192.
91. Gedik O, Akalin S, 1986 Effects of vitamin D deficiency and repletion on insulin and glucagon secretion in man. Diabetologia 29: 142-145.
92. Nyomba BL, Auwerx J, Bormans V, 1986 Pancreatic secretion in man with subclinical vitamin D deficiency. Diabetologia 29: 34-38.
93. Witham MD, Dove FJ, Dryburgh M, Sugden JA, Morris AD, Struthers AD, 2010 The effect of different doses of vitamin D(3) on markers of vascular health in patients with type 2 diabetes: a randomised controlled trial. Diabetologia 53: 2112-2119.
94. Jorde R, Sneve M, Torjesen P, Figenschau Y, 2010 No improvement in cardiovascular risk factors in overweight and obese subjects after supplementation with vitamin D3 for 1 year. J Intern 267: 462-472.
95. Patel P, Poretsky L, Liao E, 2010 Lack of effect of subtherapeutic vitamin D treatment on glycemic and lipid parameters in Type 2 diabetes: A pilot prospective randomized trial. J Diabetes 2: 36-40.
96. Parekh D, Sarathi V, Shivane VK, Bandgar TR, Menon PS, Shah NS, 2010 Pilot study to evaluate the effect of short-term improvement in vitamin D status on glucose tolerance in patients with type 2 diabetes mellitus. Endocr Pract 16: 600-608.
97. Avenell A, Cook JA, Maclennan GS, Macpherson GC, RECORD trial group, 2009 Vitamin D supplementation and type 2 diabetes: a substudy of a randomized placebo-controlled trial in older people (RECORD trial, ISRCTN 51647438). Age Ageing 38: 606-609.
98. Jorde R, Figenschau Y, 2009 Supplementation with cholecalciferol does not improve glycaemic control in diabetic subjects with normal serum 25-hydroxyvitamin D levels. Eur J Nutr 48: 349-354.
99. Nagpal J, Pande JN, Bhartia A, 2009 A double-blind, randomized, placebo-controlled trial of the short-term effect of vitamin D3 supplementation on insulin sensitivity in apparently healthy, middle-aged, centrally obese men. Diabet Med 26: 19-27.
100. Zittermann A, Frisch S, Berthold HK, et al, 2009 Vitamin D supplementation enhances the beneficial effects of weight loss on cardiovascular disease risk markers. Am J Clin Nutr 89: 1321-1327.
101. de Boer IH, Tinker LF, Connelly S, et al, 2008 Women’s Health Initiative Investigators. Calcium plus vitamin D supplementation and the risk of incident diabetes in the Women’s Health Initiative. Diabetes Care 31: 701-707.
102. Tai K, Need AG, Horowitz M, Chapman IM, 2008 Glucose tolerance and vitamin D: effects of treating vitamin D deficiency. Nutrition 24: 950-956.
103. Sugden JA, Davies JI, Witham MD, et al, 2008 Vitamin D improves endothelial function in patients with Type 2 diabetes mellitus and low vitamin D levels. Diabet Med 25: 320-325.
104. Fliser D, Stefanski A, Franek E, Fode P, Gudarzi A, Ritz E, 1997 No effect of calcitriol on insulin-mediated glucose uptake in healthy subjects. Eur J Clin Invest 27: 629-633.
105. Zofková I, Stolba P, 1990 Effect of calcitriol and trifluoperazine on glucose stimulated B cell function in healthy humans. Exp Clin Endocrinol 95: 185-191.
106. Lind L, Pollare T, Hvarfner A, Lithell H, Sørensen OH, Ljunghall S, 1989 Long-term treatment with active vitamin D (alphacalcidol) in middle-aged men with impaired glucose tolerance. Effects on insulin secretion and sensitivity, glucose tolerance and blood pressure. Diabetes Res 11: 141-147.
107. Ljunghall S, Lind L, Lithell H, et al, 1987 Treatment with one-alpha-hydroxycholecalciferol in middle-aged men with impaired glucose tolerance. A prospective randomized double-blind study. Acta Med Scand 222: 361-367.
108. Nilas L, Christiansen C, 1984 Treatment with vitamin D or its analogues does not change body weight or blood glucose level in postmenopausal women. Int J Obes 8: 407-411.
109. Harinarayan CV, Arvind S, Joshi S, Thennarasu K, Vedavyas V, Baindur A, 2014 Improvement in Pancreatic β Cell Function with Vitamin D and Calcium Supplementation in Vitamin D Deficient Non-Diabetic Subjects. Endocr Pract 20: 129-137.
Address for correspondence:
Prof. Dr. C.V. Harinarayan, M.D. (Int. Med), D.M. (Endocrinology) (AIIMS), F.A.M.S., F.R.C.P. (Glasgow), F.R.C.P. (Edinburg), Director, Institute of Endocrinology, Diabetes, Thyroid and Osteoporosis Disorders, Sakra World Hospitals, Sy No 52/2 & 53/3, Deverabeesanahalli, (opp Intel, Outer Ring Road,) Varathur Hobili, Marathahalli, Bangalore 560 103, India,E-mail: cvhari5endo@rediffmail.com
Received: 09-09-2013, Accepted: 10-01-2014