Abstract
OBJECTIVE: Stromal cell-derived factor-1 (SDF-1) is expressed in pre-adipocytes but its role is unknown. We investigated butyrate (a histone deacetylase inhibitor – HDACi) and other short-chain fatty acids (SCFA) in the regulation of SDF-1. We further investigated whether effects of SCFA were signalled through G protein-coupled receptors FFA2 and FFA3.
DESIGN AND RESULTS: SDF-1 mRNA expression and protein secretion were studied in 3T3-L1 cells and human pre-adipocytes. SDF-1 was abundant, with mRNA and protein levels increased by butyrate. This was replicated with acetate and propionate, but not with trichostatin or valproate. Trichostatin inhibited SDF-1 secretion. Pertussis toxin blocked stimulation by butyrate. The order of potency of SCFA in stimulating SDF-1 (C3 > C4 > C2) is consistent with action through FFA3. Silencing the FFA3 gene abolished butyrate-stimulated SDF-1 expression and secretion. FFA3 was expressed in both pre-adipocytes and adipocytes, while FFA2 was expressed in adipocytes only. SDF-1 expression was low in murine macrophage J774.2 cells, while the SDF-1 receptor CXCR4 was absent from 3T3-L1 cells but abundant in J774.2 macrophages. In human pre-adipocytes, FFA3 was also expressed and SCFA increased SDF-1 secretion.
CONCLUSIONS: SDF-1 and CXCR4 may mediate the interaction between adipose stromal cells and macrophages. Effects of SCFA are mediated through FFA3, but not histone deacetylase inhibition.
INTRODUCTION
Interest in the application of fatty acids as signalling molecules has increasingly grown since the recognition of a series of G protein-coupled receptors with fatty acids as their ligands.1,2 Different receptors are known to have specificities for fatty acids of differing chain length.1 Department of 7TMR Assay Development and Compound Profiling (ADCP FFA2 (GPR43) and FFA3 (GPR41) are receptors for short-chain fatty acids (SCFA) and represent a crucial part of the body’s nutrient-sensing apparatus.2 Activation of FFA2 has been reported to increase incretin secretion by enteroendocrine cells, to enhance adipogenesis, to increase leptin secretion by adipocytes, and to inhibit lipolysis.3-6 It is well-established that FFA2 is highly expressed in adipose tissue4 but may not be involved in the pathogenesis of human obesity.7 The role and expression of FFA3 is less clearly defined. Debate continues over FFA3 expression in adipose tissue, with some groups reporting no expression,3,5 while others report robust FFA3 expression in both adipose tissue and in differentiated Ob-luc adipocytes.8 Xiong et al8 also show that SCFA, acting through FFA3, increases leptin secretion and inhibits lipolysis. Taken together, these studies indicate considerable uncertainty as to the relative roles of FFA2 and FFA3 in the pathogenesis of obesity and its complications.
Interaction between cells of adipocyte lineage and those derived from bone marrow is critical in the angiogenesis accompanying adipose tissue remodelling.9 It is also important in the low-grade inflammation associated with obesity,10,11 which predisposes to insulin resistance and atherogenesis.12,13 These processes are known to be influenced by SCFA, produced in the gut by fermentation of complex carbohydrates.14-16 Stromal cell-derived factor-1 (SDF-1, CXCL12) is a chemokine which, through interaction with its obligate receptor CXCR4, is involved in angiogenesis, stem cell trafficking, and tumour metastasis.17-19 The role of the SDF-1/CXCR4 axis in adipose tissue biology has not been extensively studied to date. Interestingly, SDF-1 expression in 3T3-L1 adipocytes has been reported in two genomic studies20,21 and one proteomic study.22 Furthermore, expression of SDF-1 has been noted in adipocyte precursors with expression diminishing during adipogenesis,23 and more recently has been shown to interact with complement-derived factors in regulating adipocyte development.24 Modulation of the SDF-1/CXCR4 axis by butyrate has long been known to modulate growth of tumour tissues.25,26
The aim of this work was to examine if SCFA could affect SDF-1 gene expression and protein secretion in pre-adipocytes and investigate the mechanisms underlying this effect. More specifically, we sought to determine the relative roles of the two SFCA receptors, FFA2 and FFA3, and the role of histone deacetylase inhibition in regulating SDF-1 expression using the murine 3T3-L1 line and primary cultures of human pre-adipocytes.
MATERIALS AND METHODS
Culture and differentiation of murine 3T3-L1 cells
3T3-L1 cells (Sigma Aldrich Pty Ltd, NSW, Australia) were seeded (approximately 20,000/well) in 24-well plates and grown to confluence in Dulbecco’s Modified Eagle Medium containing 25mM glucose, bovine serum, 4mM L-glutamine, 100U/mL penicillin, and 100µg/mL streptomycin (DMEM) supplemented with 10% foetal bovine serum (FBS) at 37°C in a 5% CO2 atmosphere. For experiments on undifferentiated cells, supplement was continued for three days after reaching cell culture confluence and then replaced with DMEM containing 10% FBS without antibiotics for 24 hours before treatment with valproate or trichostatin. For experiments on differentiated cells, once pre-adipocytes were confluent, the medium was replaced with DMEM containing 10% FBS plus penicillin and streptomycin for three days. Differentiation was then initiated using 0.5mM IBMX, 1µM dexamethasone, and 10µg/ml insulin in high glucose DMEM supplemented with 10% FBS containing antibiotics (IBMX medium) for 48 hours. The cells were then maintained in DMEM supplemented with 10% FBS, insulin, and antibiotics until fully differentiated (typically 8-10 days). Prior to treatment with trichostatin or valproate, the cells were supplemented with DMEM medium with 10% FBS without antibiotics for 24 hours.
Culture of human pre-adipocytes
Approvals for the use of human tissues in this study were obtained from Queensland Health and James Cook University Human Research Ethics Committees. Adipose tissue was obtained from consenting adult patients following elective liposuction. 6 ml of adipose tissue was digested with 0.1% collagenase in PBS/BSA and placed in an orbital shaker for 60 minutes at 37°C. The solution was then filtered through a mesh (63µm) into a 50mL Falcon tube and centrifuged at 200g for 10 minutes. With the supernatant discarded, the re-suspended pellet was treated with erythrocyte lysing buffer, incubated for 5 minutes at 37°C and centrifuged at 200g for 5 minutes. Cells were counted in inoculation medium (basal medium: DMEM (50%), Ham’s F-12 (50%), HEPES, NaHCO3, biotin, and pantothenate supplemented with 10% FBS and 0.5% gentamicin) and seeded at a density of 150,000 cells per ml. After 16 hours culture, the cells were washed with Dulbecco’s Phosphate Buffered Saline (DPBS) and replaced with stimulation medium (basal medium with human transferrin, insulin, cortisol and fibroblast growth factor) until confluence was reached. To obtain adipocytes, the cells were incubated with adipogenic medium (basal medium with transferrin, gentamicin, triiodothyronine, insulin, and hydrocortisone). The medium was changed regularly until the cells differentiated. The cells were then stained for oil droplets.
Experiments on confluent 3T3-L1 pre-adipocytes and differentiated adipocytes
Treatments were prepared in DMEM supplemented with 10% FBS. Cells were washed with DPBS prior to addition of each treatment. 1ml of treatment medium was added to each well and cells incubated at 37°C, 5% CO2 for the appropriate time period. Cell medium was collected and stored at -80°C for protein expression studies. Cells were washed with DPBS (2 x 1ml) and 200µl of trypsin solution added to each well. After 5 minutes incubation, the trypsin solution was neutralized with 1ml of DMEM containing 10% FBS. The solution containing the cells was centrifuged at 460g for 5 minutes at 21°C. Medium was removed and cells were stored at -80°C pending RNA extraction.
Oil red O staining and FFA3 immunocytochemistry
Staining of accumulated lipid was performed according to kit instructions. Briefly, medium was removed from cells that were then washed with DPBS. 0.5ml Oil red O staining solution (Chemicon Australia Pty Ltd, Vic, Australia) was added to cells cultured in a 24-well plate and incubated for 15 minutes at room temperature. Staining solution was removed and cells washed with wash solution. Immunocytochemistry was used to evaluate FFA3 and FFA2 expression on THP-1 monocytes and human cultured pre-adipocytes. This was performed using rabbit polyclonal antibodies to FFA3 and FFA2 (Sapphire Biosciences, Redfern, NSW, Australia). THP-1 cells: 1×105 monocytes in 200µL medium were cytospun onto sialinised coverslips (Cytospin 4 centrifuge, Thermo Fischer Scientific, Scoresby, Vic, Australia) for 5 minutes at 500rpm. Coverslips were then air-dried and fixed in 75% ethanol and DPBS for 10 minutes.
Human pre-adipocytes cultured on slides were rehydrated with DPBS and incubated with 0.5% H2O2 to quench endogenous peroxidase activity for 10 minutes, then washed with Tris-buffered Saline with 0.05% Tween-20 (TBST buffer). This was followed by Tris Neutral Buffer (TNB) for 30 minutes and subsequently by rabbit polyclonal FFA3 antibody or isotype control (45 minutes), biotinylated rabbit IgG-G (30 minutes), HRP-streptavidin (30 minutes) with TBST wash between each step. We then added 3,3’-diaminobenzidine (DAB) chromogen prior to dehydration using an alcohol and xylene series, after staining the nucleus with Mayer’s haematoxylin. Digital images were obtained using an Olympus CKX41 microscope.
Small interfering RNA (siRNA) knockdown of FFA3
3T3-L1 pre-adipocytes were plated (20,000 cells/well) on a 6-well plate in high glucose DMEM, containing L-glutamine, penicillin, streptomycin, and supplemented with 10% FBS. The cells were incubated at 37°C in a 5% CO2 atmosphere and grown to 80% confluence. The medium was replaced with DMEM containing glutamine but without serum and antibiotics (2ml/well). The siRNA complex was formed with four separate double-stranded siRNAs designed to silence FFA3 (Qiagen Pty Ltd, Vic, Australia):
CAGCCTGGAACTGAAGGTAAA,
CAGGCTGGTCTGGTCAGTGTA and
AAGCTTCTTTCTTGGCAATTA
HiPerfect transfection reagent (Qiagen Pty Ltd, Vic, Australia) and DMEM (with no serum or antibiotics) was added and allowed to incubate for 10 minutes at 26°C. The siRNA complex was then added drop-wise onto cells (115µl/well) and mixed by gentle swirling, giving a final concentration of 5nM for each siRNA. The cells were incubated for 24 hours with the siRNA complex at 37°C, 5% CO2. The medium was then changed to include 10% FBS serum supplement and the cells were further incubated for 24 hours prior to treatment. Gene knock-down was evaluated by RT-PCR. We achieved a consistent knock-down of 75-80% (data not shown). FFA3 knock-down cells and control cells were treated with and without 2mM sodium butyrate in high glucose DMEM containing 10% FBS and L-glutamine for 24 hours. After this time, medium and cells were removed and stored at -80°C for further protein analysis.
Real-time RT-PCR analysis of mRNA expression
Total RNA was extracted from frozen (-80°C) undifferentiated and differentiated cell samples using RNeasy extraction kits (Qiagen Pty Ltd, Vic, Australia). Samples were treated with DNase as per instructions. mRNA was quantified using a Nanodrop spectrophotometer (Nanodrop Technologies, USA). Real-time RT-PCR was performed on a Corbett Rotor Gene 6 using SYBR Green and ROX. The reactions were performed in a 15µl volume containing 38ng of the extracted RNAs, 7.5µl of RT-PCR Master Mix, 0.15µl of RT Mix, and 0.6µl of the appropriate primers (SDF-1, FFA3, FFA2, PPIA, MAPK, or Pref-1). Primers were from Jomar Diagnostics Pty Ltd, SA, Australia. The reactions were normalised against peptidylpropyl isomerase A (PPI). CT values were plotted for each treatment group to determine the differences in expression level between groups.
ELISA for SDF-1
SDF-1 protein secretion was analysed using Quantikine ELISA immunoassay (Bioscientific Pty Ltd, NSW, Australia) according to manufacturer’s instructions. Cell culture supernatant (conditioned medium) was centrifuged at 12,000g for 15 minutes at 4°C. Addition of triplicate samples to the pre-coated SDF-1 plate was followed by 2 hours incubation at room temperature on an orbital shaker. The samples were then washed, followed by addition of SDF-1α HRP-conjugate (2 hours) and a subsequent wash. Substrate solution was added and the plate incubated for 30 minutes at room temperature. Stop solution was then added and optical density measured within 30 minutes at 540 nm using Tecan SunriseTM (Tecan Trading AG, Switzerland) plate reader at 450nm without correction.
Statistical Analysis
RT-PCR and ELISA data were analysed using Graphpad Prism™ software. Data were checked for normality using the Anderson-Darling Normality test. Based on the outcome of the test, parametric or non-parametric tests were carried out. Statistical comparisons between treatments were calculated using one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test corrected for multiple comparisons. Unpaired student t-test was used for direct comparison of two sets of data. Results are expressed as mean ± standard error of the mean (SEM) normalised as percentages of control unless otherwise stated. p <0.05 was deemed to be statistically significant.
RESULTS
Butyrate increases SDF-1 expression and protein secretion by 3T3-L1 cells
As expected, exposure of 3T3-L1 pre-adipocytes to IBMX medium led to cell differentiation, as evidenced by decreased expression of the pre-adipocyte marker Pref-1 (Figure 1A, p=0.002) and considerable accumulation of lipid droplets (Oil red O staining, Figure 1B). SDF-1 mRNA was abundantly expressed in pre-adipocytes and its expression was significantly increased in the presence of butyrate in a time-dependent manner (Figure 1C, p<0.001). In replicate experiments, SDF-1 mRNA was increased up to 6-fold over 24 hours. Maximal stimulation was obtained with 2mM butyrate. There was no detectable stimulation at 0.2mM nor once concentrations reached 10mM (data not shown). Butyrate also increased SDF-1 protein secretion from 3T3-L1 cells by 2.4-fold over 24 hours (Figure 1D, p <0.001). SDF-1 secretion was also apparent from differentiated 3T3-L1 cells, but there was no detected increase in secretion following exposure to butyrate.
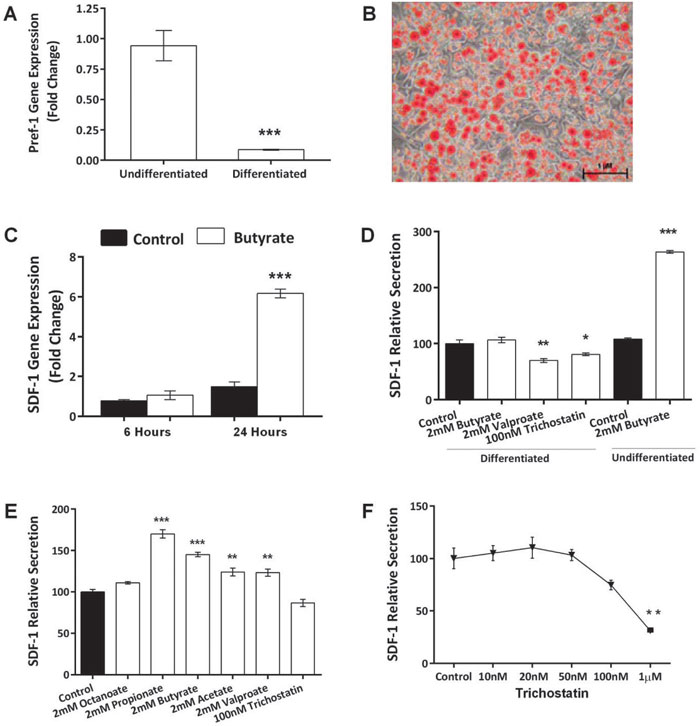
Figure 1. Butyrate increases SDF-1 expression and protein secretion by 3T3-L1 cells. A: Pref-1 expression in 3T3-L1 adipocytes. 3T3-L1 pre-adipocytes expressed high levels of Pref-1. Expression was markedly decreased when the cells were differentiated (t-test, p <0.001). B: Differentiated 3T3-L1 adipocytes, showing numerous lipid droplets stained with Oil red O. Photograph was taken 10 days after exposure of 3T3 cells to IDX medium. C: Butyrate induced a marked increase in SDF-1 mRNA determined when cells were incubated with 2 mM butyrate (ANOVA, p <0.001; control vs Butyrate 24 Hrs, p <0.001). D: Effect of HDACi (ANOVA: p <0.0001): In adipocytes, trichostatin and valproate decreased secretion of SDF-1, while butyrate was without effect. This contrasts with the stimulatory effect of butyrate in pre-adipocytes. E: Comparison of SCFA and HDACi on pre-adipocytes (ANOVA, p <0.001): Order of potency of SCFA was C3 > C4 > C2. HDACi were without effect. F: Dose-response curve for the inhibitory effect of trichostatin on SDF-1 secretion from adipocytes (ANOVA, p <0.001). * = p <0.05; ** = p <0.01; *** = p <0.001. Data shown are the mean ± SEM normalised as percentage of control. Differentiated cells =mature adipocytes; Undifferentiated cells = pre-adipocytes.
The effect of butyrate is not mediated through HDAC inhibition
Both sodium valproate (2mM) and the specific HDACi, trichostatin, modestly decreased SDF-1 secretion from differentiated 3T3-L1 cells (Figure 1D, p=0.001 and p=0.047, respectively). Compared with butyrate, there was enhanced SDF-1 secretion from 3T3-L1 pre-adipocytes in the presence of acetate (2mM, p=0.002). Propionate (2mM, p<0.001) was even more potent than butyrate (Figure 1E). The order of potency of the three SCFAs was propionate (C3) > butyrate (C4) > acetate (C2), consistent with an action through FFA3. Trichostatin decreased the expression of SDF-1 at concentrations as low as 1µM (Figure 1F, p=0.003).
The effect of butyrate is mediated through the FFA3 receptor
Pertussis toxin (PTX), which blocks GPR signalling, was used to investigate the potential role of GPR in mediating the above stimulatory effect of butyrate. When used alone at concentrations of 5nM, PTX had no effect on SDF-1 secretion by 3T3-L1 pre-adipocytes (Figure 2A, p=0.009). When added along with butyrate, PTX markedly decreased the stimulatory effect of the former on SDF-1 secretion. As there are two receptors for SCFA (FFA2 and FFA3), we investigated the relative expression of these receptors in pre-adipocytes and adipocytes using RT-PCR. FFA3 was expressed in both pre-adipocytes and in adipocytes (Figure 2B), while FFA2 was only expressed in differentiated adipocytes (Figure 2C).
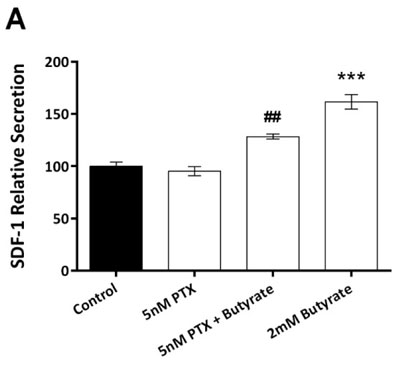
Figure 2. Pertussis toxin (PTX) abolishes the stimulatory effects of butyrate, and FFA2/FFA3 expression. A: PTX alone was without effect on SDF-1 secretion from pre-adipocytes (ANOVA, p <0.001). The stimulatory effect of butyrate was decreased in the presence of 5nM (## = p <0.01, compared against 2M Butyrate) PTX. Data shown are the mean ± SEM normalised as percentage of control. B + C: Shows 2% agarose gel electrophoresis and ethidium bromide staining. B: Expression of FFA3 (188 bp). L = ladder. Lanes 1-3 = 3T3-L1 are pre-adipocytes. Lanes 4-6 are 3T3-L1 adipocytes. N = no reverse-transcriptase negative control. C: Expression of FFA2 (156bp) – no expression was found in pre-adipocytes (lanes 1-3), while the housekeeping gene PPIA (200bp) was strongly expressed. FFA2 was expressed in differentiated adipocytes.
Using siRNA for FFA3, we successfully decreased gene expression to less than 30% of its baseline value in 3T3-L1 pre-adipocytes (Figure 3A, p=0.012) While the stimulatory effect of butyrate on SDF-1 mRNA was apparent in control cells, this effect was entirely lost in cells treated with siRNA for FFA3 (Figure 3B). Similarly, although butyrate increased SDF-1 secretion from pre-adipocytes, there was no such stimulation in cells in which the FFA3 gene had been silenced (Figure 3C).
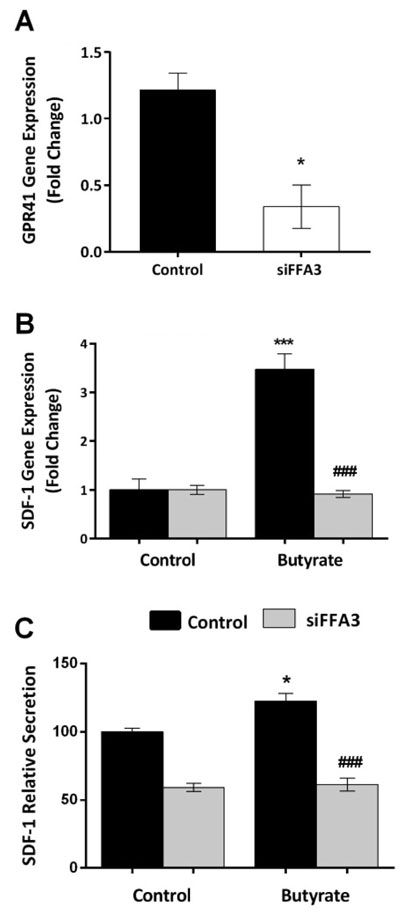
Figure 3. siRNA for FFA3 abolishes the stimulatory effect of butyrate. A: siFFA3 gene knockdown, evaluated by RT-PCR reached 75-80% (t-test, p <0.05). Data shown are the mean + SEM. B: SDF-1 m-RNA expression increased when cells were treated with butyrate (ANOVA, p<0.001, *** = p <0.001 compared with control). This stimulatory effect was markedly decreased when the FFA3 gene was silenced (### = p <0.001 compared with butyrate treatment of control cells). C:SDF-1 protein secretion was increased when control cells were treated with butyrate (ANOVA, p <0.001, * = p <0.05). This stimulation was markedly decreased following FFA3 gene silencing (### = p <0.001). Data shown are the mean ± SEM normalised as percentage of control (for B& C).
SDF-1 is a potential mediator of the interaction between pre-adipocytes and macrophages
We determined the level of SDF-1 expression and its obligate receptor (CXCR4) on cells of the mouse monocyte/macrophage line J774.2 compared with 3T3-L1 pre-adipocytes when treated with SCFA propionate (C3). Propionate was chosen as it elicited the highest SDF-1 secretion amongst the SCFAs (Figure 1E). SDF-1 protein secretion was higher in pre-adipocytes when treated with propionate compared with controls (Figure 4A, p=0.017). In contrast, SDF-1 secretion from J774.2 cells was very low and there was no significant stimulation with propionate (Figure 4A). 3T3-L1 pre-adipocytes expressed abundant SDF-1 mRNA, while there were much lower levels of expression in J774.2 cells (Figure 4B). On the other hand, CXCR4 expression was not detected in pre-adipocytes, but mRNA for CXCR4 was abundant in J774.2 cells (Figure 4C).
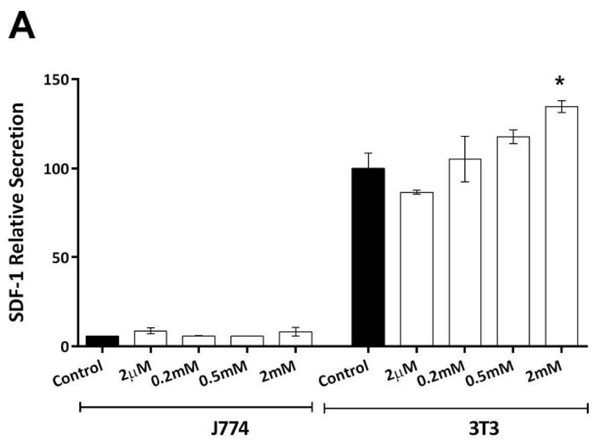
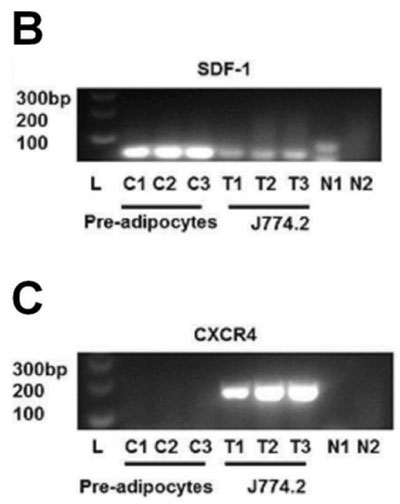
Figure 4. Effect of Propionate on SDF-1 secretion. A: Propionate enhanced SDF-1 protein secretion from pre-adipocytes, but there was only minimal SDF-1 secretion and no effect of propionate in cultures of J774.2 cells (ANOVA, p <0.001). Data shown are the mean + SEM normalised as percentage of control. * = p <0.05. B + C: 2% Agarose gel electrophoresis with ethidium bromide staining. L = ladder, N1 = no template negative control, N2 = no reverse-transcriptase negative control. SDF-1 (57bp) was detected in pre-adipocytes but not highly expressed in J774.2 cells. CXCR4 (156bp) expression was not detected in pre-adipocytes, but mRNA for CXCR4 was abundant in J774.2 cells.
Human pre-adipocytes also express SDF-1 and the FFA3 receptor
It is beyond the scope of this study to replicate all of the above work with human cells. However, SDF-1 mRNA expression was noted in human pre-adipocytes, and there was a significant increase in expression when cells were incubated in the presence of 2mM butyrate (Figure 5A, p=0.034). Using immunocytochemistry, we detected expression of FFA3, but not FFA2, in cultured human pre-adipocytes (Figure 5B-C). Both FFA2 and FFA3 were expressed on cells of the human monocyte/macrophage line THP-1 (Figure 5D-E).
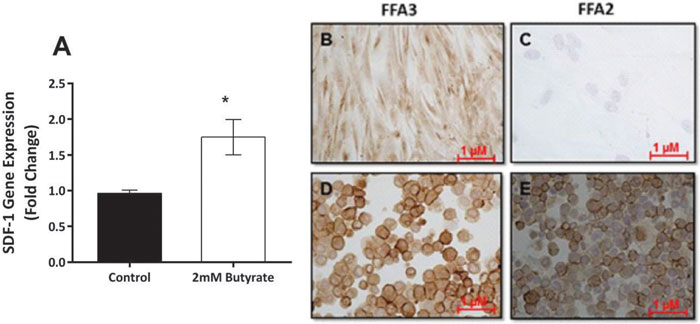
Figure 5. Human pre-adipocytes express SDF-1 and FFA3. A: SDF-1 mRNA expression was abundant in human pre-adipocytes, and this expression increased when cells were incubated with 2 mM butyrate (t-test, * = p <0.05).B: Expression of FFA3 in human pre-adipocytes. C: No expression of FFA2 in human pre-adipocytes. D: Expression of FFA3 in cells of the human monocyte/macrophage line THP-1. E: Expression of FFA2 in THP-1 cells.
DISCUSSION
In this study, we have shown that SDF-1 (CXCL12) is expressed in, and secreted by, 3T3-L1 cells. This expression is particularly evident in the pre-adipocyte state. We have also shown that SDF-1 mRNA and protein are increased by treatment with SCFA. The effect of SCFA was not replicated with the HDACs valproate and trichostatin and the inhibitory effect of pertussis toxin suggests that SCFA effects are mediated through a G protein-coupled receptor. The order of potency of SCFA (C3 > C4 > C2) suggests an effect through FFA3 rather than FFA2. Indeed FFA3, but not FFA2, was expressed in pre-adipocytes. Silencing of the FFA3 gene abolished the stimulatory effect of SCFA.
Recent studies show that SDF-1 plays a central role in the pathogenesis of diabetic retinopathy.27,28 SDF-1, through interaction with the CXCR4 receptor on bone marrow-derived vascular progenitor cells, is also crucial in vascular modelling after ischaemia.17,29,30 Decreased SDF-1 secretion in patients with diabetes is also understood to contribute to defective wound healing, including in diabetic foot ulcers.31 It is likely that cells of the stromal vascular compartment of adipose tissue are a major source of SDF-1.9,32 In obesity, these cells may be depleted due to differentiation of pre-adipocytes into mature fat-laden cells. The predominance of differentiated and hypertrophied adipocytes leads to secretion of mediators that contribute to systemic inflammation and insulin resistance.33 Our finding that pre-adipocytes are a source of SDF-1 confirms earlier observations that the chemokine is expressed in these cells.20-22
The HDACs are a group of around 18 enzymes involved in epigenetic regulation of gene expression. HDAC inhibitors are considered potential treatments for neoplasia, and neurodegenerative disorders, and as angiogenesis inhibitors. Butyrate is a naturally occurring HDAC inhibitor. While HDACs are involved in adipogenesis,34,35 the effects of HDAC inhibitors in this process remain uncertain. Valproate has been reported to inhibit adipogenesis,36 while diallyl disulphide, contained in garlic, may stimulate adipogenesis.37 SCFA have been reported to promote adipocyte differentiation and expression of adipogenic markers.5,38,39 In this study, we found that the potent HDAC inhibitor, trichostatin, inhibited SDF-1 expression in pre-adipocytes. Conversely, butyrate increased SDF-1 expression and secretion. The stimulatory effect of butyrate on SDF-1 expression and secretion was observed with other SCFAs that have only weak HDAC inhibitory activity.
In mice, plasma leptin concentration was increased by oral administration of propionate.8 The circulating propionate concentration achieved was comparable to what we used in vitro. Most of the butyrate generated within the colon is removed in the enterohepatic circulation, while high levels of acetate and propionate can reach the systemic circulation. Propionate levels are particularly high after consumption of certain functional foods, including barley kernels,40 and acetate levels also increase after alcohol consumption.41 We do not, therefore, feel that the concentrations of SCFA used in our experiments were unduly high. Additionally, we do not know what the most potent naturally occurring ligands for FFA3 are.
Given their role as part of the body’s nutrient sensing apparatus, it is not surprising to find receptors for SCFA expressed in cells of adipocyte lineage. Our results disagree with those of Hong et al3 who did not find FFA3 expression in mouse adipose depots. They did report increased adipogenesis with acetate and propionate acting through FFA2. We, in contrast, found FFA3 to be expressed in both pre-adipocytes and adipocytes, while FFA2 was only expressed in adipocytes. Xiong et al8 described the role of FFA3 in adipocyte leptin secretion induced by SCFAs. We demonstrate here that the effect of SCFA on SDF-1 was abolished with pertussis toxin, suggesting that it is GPR-mediated. We then examined the role of FFA3 in regulating SDF-1 expression. FFA3 gene silencing abolished the stimulatory effect of butyrate on SDF-1 expression and protein secretion. We also confirmed that SCFA increase SDF-1 expression in human pre-adipocytes and that FFA3, but not FFA2, is expressed in these cells.
The biological actions of SDF-1 are through its obligate receptor CXCR4 (also a co-receptor for lymphotrophic strains of the HIV virus). We did not find CXCR4 to be expressed in either pre-adipocytes or adipocytes, making it unlikely that SDF-1 and CXCR4 interaction is involved in cross-talk between mature adipocytes and stromal cells. Others have reported CXCR4 expression in adipocytes, allowing them to be infected with HIV,42 a finding not confirmed by a subsequent study.43 As expected, we detected abundant expression of CXCR4 in mouse J774.2 macrophages, but there was very little SDF-1 expression in these cells, and no effect of added propionate on SDF-1 expression. It seems highly likely therefore that SDF-1 and CXCR4 are involved in the interaction between adipocytes and macrophages, although somewhat surprisingly this has not been studied to date.
We propose that the SDF-1 secreted by adipocytes and its interaction with CXCR4 receptor on tissue macrophages mediates interaction between the two cell types (Figure 6). SDF-1 secretion by pre-adipocytes may increase monocyte infiltration into adipose and factors secreted by monocytes may inhibit adipocyte differentiation.10,44 In states of fat excess, such impaired differentiation may lead to ectopic deposition of fat, including in the vascular wall. On the other hand, hypertrophied33 or dead45 adipocytes attract macrophages and such infiltration in visceral fat partly accounts for the association between visceral obesity and systemic inflammation and insulin resistance.46,47
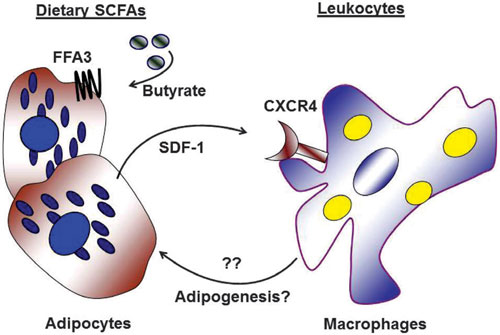
Figure 6. Hypothetical adipocyte-macrophage interactions. Dietary SCFAs including butyrate induce adipocytes to secrete SDF-1, acting through the FFA3 receptor. Secreted SDF-1 attracts other cells including macrophages, interacting through its receptor CXCR-4. This adipocyte-macrophage interaction could initiate secretion of mediators from macrophages, possibly regulating processes associated with adipogenesis and insulin resistance.
CONCLUSIONS
In conclusion, we have demonstrated that SDF-1 (CXCL12) is secreted by 3T3-L1 pre-adipocytes and adipocytes and that this is regulated by SCFA in a mechanism that does not depend on histone deacetylase inhibition. Our data strongly suggest that the effect of SCFA is mediated through FFA3. SDF-1 may be an important mediator of the interaction between adipocytes and bone marrow-derived cells including monocyte/macrophage cells. These observations shed light on a novel pathway mediating interaction between adipocyte precursors and inflammatory cells. Further studies may increase our understanding of the processes involved in adipose depot remodelling and the systemic inflammation that accompanies obesity.
AKNOWLEDGEMENTS
We are grateful to Frances Wood, Monsur Kazi, and Stephen Garland for their technical expertise and contribution to this work.
We would like also to thank Dr. Mark Vucak, Consultant Plastic Surgeon, Mater Hospital, Townsville, who kindly supplied the samples from which human pre-adipocytes were cultured.
FUNDING
We appreciate the support of James Cook University and the Private Practice Fund of the Townsville Hospital, Queensland, Australia for funding this research.
REFERENCES
1. Brown AJ, Jupe S, Briscoe CP, 2005 A family of fatty acid binding receptors. DNA Cell Biol 24: 54-61.
2. Covington DK, Briscoe CA, Brown AJ, Jayawickreme CK, 2006 The G-protein-coupled receptor 40 family (GPR40-GPR43) and its role in nutrient sensing. Biochem Soc Trans 34: 770-773.
3. Hong YH, Nishimura Y, Hishikawa D, et al, 2005 Acetate and propionate short chain fatty acids stimulate adipogenesis via GPCR43. Endocrinology 146: 5092-5099.
4. Ge H, Li X, Weiszmann J, et al, 2008 Activation of G protein-coupled receptor 43 in adipocytes leads to inhibition of lipolysis and suppression of plasma free fatty acids. Endocrinology 149: 4519-4526.
5. Zaibi MS, Stocker CJ, O’Dowd J, et al, 2010 Roles of GPR41 and GPR43 in leptin secretory responses of murine adipocytes to short chain fatty acids. FEBS Lett 584: 2381-2386.
6. Tolhurst G, Heffron H, Lam YS, et al, 2012 Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes 61: 364-371.
7. Dewulf EM, Ge Q, Bindels LB, et al, 2013 Evaluation of the relationship between GPR43 and adiposity in human. Nutr Metab (Lond) 10: 11.
8. Xiong Y, Miyamoto N, Shibata K, et al, 2004 Short-chain fatty acids stimulate leptin production in adipocytes through the G protein-coupled receptor GPR41. Proc Natl Acad Sci U S A 101: 1045-1050.
9. Cho CH, Koh YJ, Han J, et al, 2007 Angiogenic role of LYVE-1-positive macrophages in adipose tissue. Circ Res 100: e47-57.
10. Lacasa D, Taleb S, Keophiphath M, Miranville A, Clement K, 2007 Macrophage-secreted factors impair human adipogenesis: involvement of proinflammatory state in preadipocytes. Endocrinology 148: 868-877.
11. Oh DY, Lagakos WS, 2011 The role of G-protein-coupled receptors in mediating the effect of fatty acids on inflammation and insulin sensitivity. Curr Opin Clin Nutr Metab Care 14: 322-327.
12. Permana PA, Menge C, Reaven PD, 2006 Macrophage-secreted factors induce adipocyte inflammation and insulin resistance. Biochem Biophys Res Commun 341: 507-514.
13. Harford KA, Reynolds CM, McGillicuddy FC, Roche HM, 2011 Fats, inflammation and insulin resistance: insights to the role of macrophage and T-cell accumulation in adipose tissue. Proc Nutr Soc 70: 408-417.
14. Maslowski KM, Vieira AT, Ng A, et al, 2009 Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 461: 1282-1286.
15. Tian J, Dang HN, Yong J, et al, 2011 Oral treatment with y-aminobutyric acid improves glucose tolerance and insulin sensitivity by inhibiting inflammation in high fat diet-fed mice. PLoS ONE 6: e25338.
16. Marsland BJ, 2012 Regulation of inflammatory responses by the commensal microbiota. Thorax 67: 93-94.
17. Ghadge SK, Muhlstedt S, Ozcelik C, Bader M, 2011 SDF-1 as a therapeutic stem cell homing factor in myocardial infarction. Pharmacol Ther 129: 97-108.
18. Liu G, Lu P, Li L, et al, 2011 Critical role of SDF-1-induced progenitor cell recruitment and macrophage VEGF production in the experimental corneal neovascularization. Mol Vis 17: 2129-2138.
19. Ji RC, 2012 Macrophages are important mediators of either tumor- or inflammation-induced lymphangiogenesis. Cell Mol Life Sci 69: 897-914.
20. Burton GR, Guan Y, Nagarajan R, McGehee RE Jr, 2002 Microarray analysis of gene expression during early adipocyte differentiation. Gene 293: 21-31.
21. Gerhold DL, Liu F, Jiang G, et al, 2002 Gene expression profile of adipocyte differentiation and its regulation by peroxisome proliferator-activated receptor-gamma agonists. Endocrinology 143: 2106-2118.
22. Kratchmarova I, Kalume DE, Blagoev B, et al, 2002 A proteomic approach for identification of secreted proteins during the differentiation of 3T3-L1 preadipocytes to adipocytes. Mol Cell Proteomics 1: 213-222.
23. Oh I, Ozaki M, Miyazato A, et al, 2007 Screening of genes responsible for differentiation of mouse mesenchymal stromal cells by DNA micro-array analysis of C3H10T1/2 and C3H10T1/2-derived cell lines. Cytotherapy 9: 80-90.
24. Blogowski W, Budkowska M, Salata D, et al, 2013 Clinical analysis of selected complement-derived molecules in human adipose tissue. J Transl Med 11: 11.
25. Jordan NJ, Kolios G, Abbot SE, et al, 1999 Expression of functional CXCR4 chemokine receptors on human colonic epithelial cells. J Clinl Invest 104: 1061-1069.
26. Shibuta K, Mori M, Shimoda K, et al, 2002 Regional expression of CXCL12/CXCR4 in liver and hepatocellular carcinoma and cell-cycle variation during in vitro differentiation. Jpn J Cancer Res 93: 789-797.
27. Butler JM, Guthrie SM, Koc M, et al, 2005 SDF-1 is both necessary and sufficient to promote proliferative retinopathy. J Clin Invest 115: 86-93.
28. Abu El-Asrar AM, Struyf S, Kangave D, Geboes K, Van Damme J, 2006 Chemokines in proliferative diabetic retinopathy and proliferative vitreoretinopathy. Eur Cytokine Netw 17: 155-165.
29. Gallagher KA, Liu ZJ, Xiao M, et al, 2007 Diabetic impairments in NO-mediated endothelial progenitor cell mobilization and homing are reversed by hyperoxia and SDF-1 alpha. J Clin Invest 117: 1249-1259.
30. van Weel V, Seghers L, de Vries MR, et al, 2007 Expression of vascular endothelial growth factor, stromal cell-derived factor-1, and CXCR4 in human limb muscle with acute and chronic ischemia. Arterioscler Thromb Vasc Biol 27: 1426-1432.
31. Di Rocco G, Gentile A, Antonini A, et al, 2010 Enhanced healing of diabetic wounds by topical administration of adipose tissue-derived stromal cells overexpressing stromal-derived factor-1: biodistribution and engraftment analysis by bioluminescent imaging. Stem Cells Int 2011: 304562.
32. Thangarajah H, Vial IN, Chang E, et al, 2009 IFATS collection: Adipose stromal cells adopt a proangiogenic phenotype under the influence of hypoxia. Stem Cells 27: 266-274.
33. Skurk T, Alberti-Huber C, Herder C, Hauner H, 2007 Relationship between adipocyte size and adipokine expression and secretion. J Clin Endocrinol Metab 92: 1023-1033.
34. Fajas L, Egler V, Reiter R, et al, 2002 The retinoblastoma-histone deacetylase 3 complex inhibits PPARgamma and adipocyte differentiation. Dev Cell 3: 903-910.
35. Li D, Yea S, Li S, et al, 2005 Kruppel-like factor-6 promotes preadipocyte differentiation through histone deacetylase 3-dependent repression of DLK1. J Biol Chem 280: 26941-26952.
36. Lagace DC, Nachtigal MW, 2004 Inhibition of histone deacetylase activity by valproic acid blocks adipogenesis. J Biol Chem 279: 18851-18860.
37. Lee JH, Kim KA, Kwon KB, et al, 2007 Diallyl disulfide accelerates adipogenesis in 3T3-L1 cells. Int J Mol Med 20: 59-64.
38. Toscani A, Soprano DR, Soprano KJ, 1990 Sodium butyrate in combination with insulin or dexamethasone can terminally differentiate actively proliferating Swiss 3T3 cells into adipocytes. J Biol Chem 265: 5722-5730.
39. Yoo EJ, Chung JJ, Choe SS, Kim KH, Kim JB, 2006 Down-regulation of histone deacetylases stimulates adipocyte differentiation. J Biol Chem 281: 6608-6615.
40. Nilsson A, Granfeldt Y, Ostman E, Preston T, Bjorck I, 2006 Effects of GI and content of indigestible carbohydrates of cereal-based evening meals on glucose tolerance at a subsequent standardised breakfast. Eur J Clin Nutr 60: 1092-1099.
41. Sarkola T, Iles MR, Kohlenberg-Mueller K, Eriksson CJ, 2002 Ethanol, acetaldehyde, acetate, and lactate levels after alcohol intake in white men and women: effect of 4-methylpyrazole. Alcohol Clin Exp Res 26: 239-245.
42. Hazan U, Romero IA, Cancello R, et al, 2002 Human adipose cells express CD4, CXCR4, and CCR5 [corrected] receptors: a new target cell type for the immunodeficiency virus-1? Faseb J 16: 1254-1256.
43. Munier S, Borjabad A, Lemaire M, Mariot V, Hazan U, 2003 In vitro infection of human primary adipose cells with HIV-1: a reassessment. Aids 17: 2537-2539.
44. Constant VA, Gagnon A, Landry A, Sorisky A, 2006 Macrophage-conditioned medium inhibits the differentiation of 3T3-L1 and human abdominal preadipocytes. Diabetologia 49: 1402-1411.
45. Cinti S, Mitchell G, Barbatelli G, et al, 2005 Adipocyte death defines macrophage localization and function in adipose tissue of obese mice and humans. J Lipid Res 46: 2347-2355.
46. Cancello R, Tordjman J, Poitou C, et al, 2006 Increased infiltration of macrophages in omental adipose tissue is associated with marked hepatic lesions in morbid human obesity. Diabetes 55: 1554-1561.
47. Okamoto Y, Higashiyama H, Rong JX, et al, 2007 Comparison of mitochondrial and macrophage content between subcutaneous and visceral fat in db/db mice. Exp Mol Pathol 83: 73-83.
Address for correspondence:
Professor R.L. Kennedy, School of Medicine, Deakin University, Waurn Ponds Campus, Geelong VIC 3220, Australia, Tel.: 03-5227-1275, Fax: 03-5246-2986, E-mail: richardlee.k@hotmail.com
Received 23-12-2013, Accepted 03-04-2014