HORMONES 2005, 4(3):155-160
DOI: 10.14310/horm.2002.——
Address correspondence and requests for reprints to:
Kiapekou Erasmia, 25 Karaiskaki Str., 15772 Athens, Greece, Tel.: +30 210 7488926, E-mail: nem10@panafonet.gr
Received 11-06-05, Revised 10-07-05, Accepted 15-07-05
Abstract
Objective: A number of hormones and growth factors have been reported to affect the in vitro maturation of oocytes. Their exact effects on follicular growth and oocyte maturation and the mechanisms involved are still unclear. In the present study, we have investigated the effects of Growth Hormone (GH) and Insulin-like Growth Factor 1 (IGF-1) on the in vitro maturation of mouse oocytes. Design: Immature preovulatory oocytes without cumulus cells (denuded), were obtained from 4- to 8-week old female mice and were cultured in Ham’s F-10 medium. GH and IGF-1 were added separately or in combination in gradually increasing concentrations in the culture media, while medium-only containing samples were employed as controls. Oocyte development was assessed daily for three days and maturation was considered to be completed when the first polar body appeared. Results: In control samples, 44±3% (mean ± SE) of denuded oocytes formed a polar body. The achieved maturation rate after the addition of either GH or IGF-1 or GH plus IGF-1 was significantly higher than in controls. The highest maturation rates were achieved after the addition of 0.2 µg/ml GH (76%±5%), 50 ng/ml IGF-1 (69%±5%) and a combination of 0.2 µg/ml GH plus 10 ng/ml IGF-1 (75%±5%). Conclusions: The data suggest that GH and IGF-1, alone or in combination, affect mouse oocyte maturation significantly. The lack of a synergistic effect on oocyte cultures when both hormones were added indicates that both hormones act through the same signaling pathway.
INTRODUCTION
In vitro maturation of oocytes is a long and complex process that has not yet been completely elucidated. Gonadotropins are considered to be of considerable importance in the maturation of oocytes. In addition, a local regulation of the ovarian function by steroid and non-steroid substances has recently been described. The latter include a variety of growth factors, such as Growth Hormone (GH)1, Insulin-like Growth Factors (IGFs)2 and the Epidermal Growth Factor (EGF)3, as well as other hormones such as Prolactin (PRL)4. These growth factors are secreted from the granulosa and cumulus cells in response to gonadotropins and subsequently act on the oocyte via paracrine and autocrine pathways. Although a growing number of studies have indicated that there are beneficial effects of growth factors in oocyte maturation, it seems that only denuded oocytes require the supplementation of growth factors in the culture medium for proper maturation5. Many of those agents are produced from the follicle itself and have either an autocrine or paracrine mechanism of action.
Recently, GH has been implicated in oocyte maturation. Izadyar et al6 reported an acceleration of in vitro oocyte maturation, of cumulus-enclosed oocytes, the induction of cumulus expansion and the promotion of subsequent embryonic development of bovine oocytes with the addition of GH in the cultures. The GH mRNA expression and GH localisation have been demonstrated in the cumulus cells and the enclosed oocyte of small to medium sized bovine follicles7. Moreover, the addition of GH appears to increase the in vitro maturation rate of early-antral follicles in the rat8, while the presence of GH receptor mRNA has been established in rat preantral follicles9. Furthermore, GH promotes preantral follicular growth in 11-day old immature female mice10. Growth hormone can act directly on rat follicles through its receptors at the granulosa cells9 and/or indirectly through IGF-18. This is also the case in porcine12 and rabbit13 follicles and denuded oocytes (oocytes at the late stages of development, not surrounded by layers of cumulus cells and the germinal vesicle clear and distinguished).
The presence of IGF-1 mRNA and IGF-1 receptor mRNA in rat somatic follicular cells and in oocytes has been demonstrated11. Furthermore, the addition of IGF-1 to the culture medium increases the in vitro nuclear maturation rate13. Studies in humans have shown that IGF-1 receptors are present in the follicles throughout all stages of development (from the primordial through to the preovulatory stage)14. In addition, IGF-1 promotes the in vitro maturation of follicles and denuded oocytes15. Interestingly, Insulin-like Growth Factor 1 enhances the stimulatory effects of GH on the in vitro maturation of mouse preantral follicles10.
The aim of the present study was to investigate the effect of GH and IGF-1, given separately or in combination, on the in vitro maturation of mouse denuded oocytes using a simple in vitro culture system.
METHODS
Animals
Denuded oocytes were obtained from 4- to 8-week old female mice without prior hormonal ovarian stimulation [New Zealand Black (NZB) x New Zealand White (NZW); (Pasteur Institute, Athens)]. The mice were sacrificed by cervical dislocation and the ovaries were removed.
Materials
Dulbecco’s phosphate-buffered saline (DBPS) and Ham’s F-10 medium without hypoxanthine were obtained from Invitrogen Life Technologies, (Paisley, UK); bovine serum albumin (BSA), from Sigma-Aldrich, (Dorset, UK); Human growth hormone from Eli Lilly (Windlesham Surrey, U.K.); Human Insulin-like Growth Factor Type 1 from R&D Systems (Minneapolis, USA); culture dishes (Falcon), from Becton Dickinson Co., (New Jersey, USA).
Isolation of mouse denuded oocytes
Mouse denuded oocytes were collected mechanically by puncturing the ovaries in Dulbecco’s phosphate-buffered saline, supplemented with bovine serum albumin (9:1). Approximately 1000 mouse denuded oocytes were obtained. Oocytes were thus isolated at the late stages of development, when not surrounded by layers of cumulus cells, and the germinal vesicle was clearly visible (Figure 1).
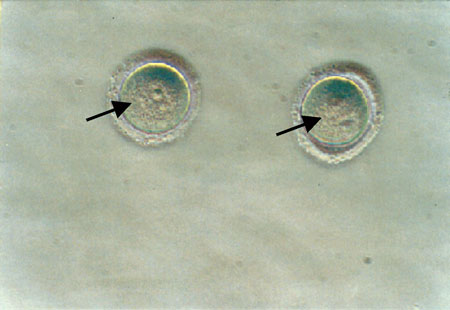
Figure 1. Oocytes at the late stages of development, not surrounded by layers of cumulus cells and the germinal vesicle clearly distinguished. The oocytes were collected from the ovaries of 2- to 8-week old mice, without hormonal ovarian stimulation. The germinal vesicles of the oocytes are visible (black arrow) (magnification x30).
Culture of mouse denuded oocytes
Collected oocytes were rinsed twice in a DPBS and BSA mixture solution (9:1) and cultured in Ham’s F-10 medium without hypoxanthine supplemented with BSA (9:1). Growth hormone and IGF-1 were added as described below, while medium-only samples were used as controls. Ten to twenty oocytes were placed in the central well of the culture dishes with 1 ml of Ham’s F-10 medium. Additionally, 4 ml of Ham’s F-10 medium were added to the surrounding area of the dish to maintain humidity.
Culture conditions were adjusted at 37oC, 5% CO2 concentration, 95% humidity and pH 7.3 – 7.4. Oocyte development was assessed daily for 72 hours for the appearance of the first polar body.
Dose-Response Experiments
Gradually increasing concentrations of human GH (0.1, 0.2, 0.4 and 0.6 µg/ml) and human IGF-1 (10, 50, 100 and 200 ng/ml) were added separately to different cultures and their effect in mouse oocyte development was recorded. Furthermore, based on the preliminary results obtained from the dose-response experiments, all tested doses of GH were added to different cultures together with 10 ng/ml IGF-1. The concentrations of GH and IGF-1 that we used have been employed and mentioned in previous publications8,11,12.
Statistics
The results were analysed by one-way analysis of variance and the z-test for proportions. A value of P <0.05 was considered statistically significant. Percentages of in vitro oocyte maturation were expressed as mean±SE. The Bonferroni correction was applied.
RESULTS
The highest maturation rate was observed within the first 24 hrs. There was a small number of mature oocytes in the second 24hr period, subsequent to which most oocytes degenerated. We observed their maturation for an additional 24hr period but at that point most oocytes had degenerated16.
Control cultures
Mouse denuded oocytes, cultured without GH or IGF-1 for 72 hours, matured at a rate of 44±%.
Effects of GH on the in vitro maturation of mouse denuded oocytes
The maturation rate of cultured mouse denuded oocytes significantly increased (p <0.05) in the presence of 0.1 µg/ml or 0.2 µg/ml of GH (mean±SE; 67±1% and 76±5%, respectively, versus 44±3% in control cultures) (Figure 2). Further increase in the GH concentrations added to the cultures (0.4 µg/ml and 0.6 µg/ml) did not result in any significant rise in the maturation rate as compared to control samples.
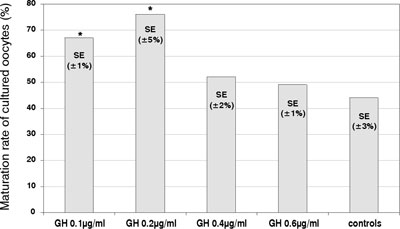
Figure 2. Effects of GH on the in vitro maturation of mouse denuded oocytes. The percentage of cultured oocytes developing a polar body after 72 hours (maturation rate), in the absence (control) or the presence of GH (µg/ml) at various concentrations. (* – significantly different from the controls; p<0.05).
Effects of IGF-1 on the in vitro maturation of mouse denuded oocytes
The maturation rate of cultured mouse denuded oocytes significantly increased (p <0.05) in the presence of 10 and 50 ng/ml of IGF-1 (mean±SE; 65±2% and 69±5%, respectively, versus 44±3% in control cultures) (Figure 3). Further increase of IGF-1 concentrations did not result in any significant rise in the maturation rate as compared to control samples.
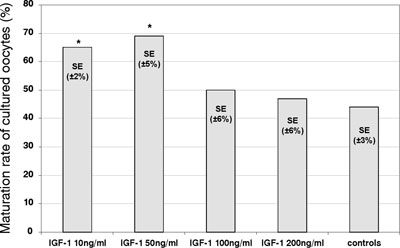
Figure 3. Effects of IGF-1 on the in vitro maturation of mouse denuded oocytes. The percentage of cultured oocytes developing a polar body after 72 hours (maturation rate), in the absence (control) or the presence of IGF-1 (ng/ml) at various concentrations. (* – significantly different from the controls; p<0.05).
Effects of GH plus IGF-1 on the in vitro maturation of mouse denuded oocytes
The maturation rate of cultured mouse denuded oocytes significantly increased (p <0.05) in the presence of 10 ng/ml plus either 0.1 µg/ml or 0.2 µg/ml of GH (mean±SE; 70±2% and 75±5%), as compared to control cultures (mean±SE; 44±3%) (Figure 4). Further increase of GH concentrations did not result in any significant changes in the maturation rate as compared to control samples.
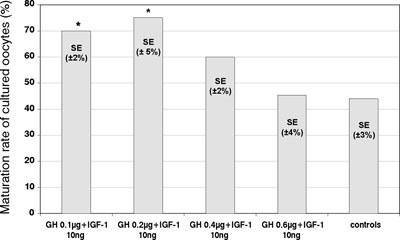
Figure 4. Effects of different combinations of GH (ìg/ml) and IGF-1 (ng/ml) on the in vitro maturation of mouse denuded oocytes. The percentage of cultured oocytes developing a polar body after 72 hours (maturation rate), in the presence of IGF-1 and GH at various concentrations and in controls (* – significantly different from the controls; p<0.05).
DISCUSSION
We found that the in vitro maturation rates of mouse denuded oocytes are significantly increased in the presence of GH and/or IGF-1.Increasing the concentrations of GH and IGF-1 beyond a certain level does not further increase oocyte maturation. This could be attributed to a down-regulation of their corresponding receptors when higher concentrations are employed. The down-regulation phenomenon is often observed when there are increased concentrations of the ligand in the culture media. It is readily seen with protein membrane receptors and represents a physiological mechanism of cellular adaptation to increased concentrations of the ligand.
Growth hormone promotes preantral follicular growth in 11-day old immature female mice10. The mode of action of GH on oocytes is not well known. Two different pathways have been suggested. The first is a direct one, through GH receptors 9, while the second is indirect, through the action of granulosa-derived IGF-112,13. In bovine oocytes, some studies showed that GH acts via IGF-117 while other studies showed that GH acts directly via a cAMP-mediated pathway18,19. Growth hormone receptor is expressed in bovine and rat cumulus cells and in oocytes18,20,21. In the past it had been suggested that the effects of GH on oocyte maturation were exerted via cumulus cells in the two species8,18. In the present study we employed mouse denuded oocytes and found that GH as well as IGF-1 significantly increased the in vitro maturation rate of these oocytes. The absence of any supporting cumulus cells suggests the presence of GH receptor on oocytes at this developmental stage in mice. Furthermore, GH and GH receptor mRNA were demonstrated to be expressed in bovine oocytes and granulosa cells recovered from small follicles maturing in vitro7. In granulosa cells, GH helps the differentiation and replication22 in addition to the modulation of steroidogenic gene expression23.
Previous studies have demonstrated the presence of IGF-1 and IGF-1 receptor mRNA at the cumulus cells and the denuded bovine and porcine oocytes24,25. In this study IGF-1 appears to act directly on the oocytes of late stage development without the presence of any cumulus cells. In mice, low IGF-1 mRNA levels were detected in primary follicles while transcription increased to maximum during the late preantral and early antral stages2,26. Studies on IGF-1 knockout mice have demonstrated the inability of the follicles to reach the antral stage, inducing infertility due to lack of ovulation27, thus confirming the essential role of IGF-1 during folliculogenesis. Demeestere et al have shown that IGF-1 has a stimulatory effect on follicular steroidogenesis if added to culture of mouse preantral follicles in the presence of FSH. Subsequently, blastocyst development was enhanced after follicular culture with IGF-128. The presence of IGF-1 receptor mRNA on rat denuded oocytes and follicles has been described11,29. In the present study, IGF-1 stimulated the in vitro maturation of mouse denuded oocytes without the presence of FSH, suggesting that the action of IGF-1 in mouse denuded oocytes is not FSH-dependent and that it might be exerted through IGF-1 receptors.
The combination of IGF-1 with various concentrations of GH on mouse cultures did not result in a higher maturation rate than that observed by the sole utilisation of either IGF-1 or GH, suggesting that the two hormones do not act synergistically, possibly because they act through the same signaling pathway.
In conclusion, our data showed that GH and/or IGF-1 increase the rate of the in vitro maturation of mice denuded oocytes. In addition, a possible relationship between their pathways of action has been proposed.
REFERENCES
1. Apa R, Lanzone A, Miceli F, et al, 1995 Growth hormone-releasing factor stimulates meiotic maturation in follicle- and cumulus-enclosed rat oocytes. Mol Cell Endocrin 112: 195-201.
2. Adashi EY, Resnick CE, Svoboda ME, Van Wyk JJ, 1988 In vivo regulation of granulose cell somatomedin- C/insulin-like growth factor-I receptors. Endocrinology 122: 1383-1390.
3. Reed M , Estrada J, Illera M, Peters R, 1993 Effects of epidermal growth factor, insulin-like growth factorI, and dialyzed porcine follicular fluid on porcine oocyte maturation in vitro. J Exp Zool 266: 74-78.
4. Yoshimura Y, Hosoi Y, Iritani A, Nakamura Y, Atlas SJ, Wallach EE, 1989 Developmental potential of rabbit oocyte matured in vitro:the possible role of prolactin. Biol Reprod 41: 26-33.
5. Chian RC, Tan SL, 2002 Maturational and developmental competence of cumulus free immature human oocytes derived from stimulated and intracytoplasmic sperm injection cycles. Reprod Biomed Online 5: 125_132.
6. Izadyar F, Colenbrander B, Bevers MM, 1996 In vitro maturation of bovine oocytes in the presence of growth hormone accelerates nuclear maturation and promotes subsequent embryonic development. Mol Reprod Dev 45: 372_377.
7. Izadyar F, Zhao J, Van Tol HTA, Colenbrander B, Bevers MM, 1999 Messenger RNA expression and protein localization of growth hormone in bovine ovarian tissue and in cumulus oocyte complexes (COCs) during in vitro maturation. Mol Reprod Dev 53: 398-406.
8. Apa R, Lanzone A, Miceli F, et al, 1994 Growth hormone induces in vitro maturation of follicle- and cumulus- enclosed rat oocytes. Mol Cell Endocrin 106: 207-212.
9. Zhao J, Van Tol HTA, Taverne MAM, Van der Weijden GC, Bevers MM, Van Den Hurk R, 2000 The effect of growth hormone on pre-antral follicles in vitro. Zygote 8: 275-283.
10. Liu X, Kazumichi A, Yokota H, et al, 1998 Effects of growth hormone, activin and follistatin on the development of preantral follicle from immature female mice. Endocrinology 139: 2342-2347.
11. Zhao J, Taverne MAM, Van Der Weijden GC, Bevers MM, Van Den Hurk R, 2001 Insulin-like growth factor-1 (IGF-1) stimulates the development of pre-antral follicles. Mol Reprod Dev 58: 287-296.
12. Sirotkin A, Taradajnik T, Makarevich A, Bulla J, 1998 Effect of follicular cells, IGF-I and tyrosine kinase blockers on oocyte maturation. Anim Reprod Science 51: 333-344.
13. Yoshimura Y, Ando M, Nagamatsu M, et al, 1996 Effects of insulin-like growth factor-I on follicle growth, oocyte maturation and ovarian stereidogenesis and plasminogen activatior activity in rabbit. Biol Reprod 55: 152-160.
14. Zhou J, Bondy C, 1993 Anatomy of the human ovarian insulin-like growth factor system. Biol Reprod 43: 467-474.
15. Gomez E, Tarin JJ, Peiiicer A, 1993 Oocyte maturation in humans:the role of gonadotropins and growth factors. Fertil Steril 60: 40-46.
16. Nakano H, Kubo H, 2000 Study of the in vitro maturation of mouse oocytes induced by microinjection of maturation promoting factor (MPF). J Assist Reprod Genet 17: 67-73.
18. Izadyar F, Van Tol HTA, Colenbrander B, Bevers MM, 1997 Stimulatory effect of growth hormone on in vitro maturation of bovine oocytes is exerted through cumulus cells and not mediated by IGF-I. Mol Reprod Dev 47: 175_180.
19. Izadyar F, Colenbrander B, Bevers MM, 1997 Stimulatory effect of growth hormone on in vitro maturation of bovine oocytes is exerted through the cyclic adenosine 3′, 5′-monophosphate signaling pathway. Biol Reprod 57: 1484_1489.
20. Lucy MC, Collier RJ, Kitchell ML, Dibner JJ, Hauser SD, Krivi GG, 1993 Immunohistochemical and nucleic acid analysis of somatotropin receptor populations in the bovine ovary. Biol Reprod 48: 1219_1227.
21. Kolle S, Sinowatz F, Boie G, Lincoln D, 1998 Developmental changes in the expression of the growth hormone receptor messenger ribonucleic acid and protein in the bovine ovary. Biol Reprod 59: 836_842.
22. Izadyar F, Hage WJ, Colenbrander B, Bevers MM, 1998 The promotory effect of growth hormone on the developmental competence of in vitro matured bovine oocytes is due to improved cytoplasmic maturation. Mol Reprod Dev 49: 444_453.
23. Doldi N, Bassan M, Bonzi V, Ferrari A, 1996 Effects of growth hormone and growth hormone-releasing hormone on steroid synthesis in cultured human luteinizing granulosa cells. Gynecol Endocrinol 10: 101-108.
24. Lorenzo PL, Illera MJ, Illera JC, Illera M, 1994 Enhasment of cumulus expansion and nuclear maturation during bovine oocyte IVM with the addition of epidermal growth factor and insulin-like growth factor I. J Reprod Fertil 101: 697-701.
25. Xia P, Tekpetey FR, Armstrong DT, 1994 Effect of IGF-I on pig oocyte maturation, fertilization, and embryonic development in vitro, and on granulose and cumulus cell biosynthetic activity. Mol Reprod Dev 38: 373-379.
26. Wandji SA, Wood TL, Crawford J, Levison SW, Hammond JM, 1998 Expression of mouse ovarian insulin growth factor system components during follicular development and atresia. Endocrinology 139: 5205-5214.
27. Baker J, Hardy MP, Zhou J, Bondy C, Lupu F, Bellve AR, Efstratiadis A, 1996 Effects of an Igf1 gene null mutation on mouse reproduction. Mol Endocrinol 10: 903-918.
28. Demeestere I, Gervy C, Centner J, Devreker F, Englert Y, Delbaere A, 2004 Effect of insulin-like growth factor-I during preantral follicular culture on steroidogenesis, in vitro oocyte maturation, and embryo development in mice.Biol Reprod 70: 1664-1669.
29. Adashi E, Resnick E, Payne D, et al, 1997 The mouse intraovarian Insulin-like Growth Factror I system: Departures from the rat paradigm. Endocrinology 138: 3881-3890.