Abstract
OBJECTIVE: In NOD.H2h4 mice, high dietary iodine intake has been known to cause Iodine-accelerated Spontaneous Autoimmune Thyroiditis (ISAT) via an unknown mechanism. The aim of the study was to examine whether the NOD.h4 genetic background predisposes to enhanced iodine organification in thyroglobulin (Tg), a target autoantigen in ISAT. DESIGN: To avoid issues associated with an ongoing anti-Tg antibody response, we assessed Tg iodination levels in iodine-fed, B-cell deficient NOD.h4 mice. Additionally, we tested whether humoral or cellular immune responses of iodine-fed NOD.h4 mice are preferentially directed to Tg with increased iodine content (I-Tg) or known pathogenic Tg peptides that contained iodine. RESULTS: The iodine content of Tg was not significantly different between control (9.0±2.7 I atoms per monomer) and iodine-fed mice (10.9±0.3 I atoms per monomer). Furthermore, in iodine-fed NOD.h4 mice developing ISAT, strong but equivalent serum IgG responses were detected to both Tg or I-Tg, whereas their lymphoid cells were stimulated weakly but equally well by Tg or I-Tg in vitro and did not show reactivity against a panel of five pathogenic Tg peptides that contained iodine. CONCLUSIONS: The results suggest that development of ISAT in NOD.h4 mice is not associated with enhanced iodine organification or differential B- or T-cell responses to iodinated determinants in Tg.
INTRODUCTION
The interplay between genetic predisposition and environment underlying the induction of autoimmune disease has been clearly shown in NOD.H2h4 mice, a model of Spontaneous Autoimmune Thyroiditis (SAT). This congenic strain, expressing the Kk, Ak, E0, Db molecules in the NOD background, develops periinsulitis but not diabetes1 and exhibits SAT at a higher incidence (50%-70% vs 5%) than the NOD strain.2,3 Addition of NaI to the drinking water accelerates the onset of SAT from 7-10 months in control mice to 2 months following the initiation of additional NaI intake in NOD.H2h4 hosts2 and significantly enhances both its incidence and severity.4,5 Increased iodine ingestion has previously shown similar effects in other animal models of SAT such as the CS chicken6 and the BB/w rat,7 but the NOD.H2h4 model has attracted attention in recent years due to the availability of well characterized mouse biomarkers and reagents. Epidemiological data clearly support the view that an iodine-rich diet can also promote subclinical hypothyroidism in humans,8-10 but the pathogenetic mechanisms remain mostly unknown.
Thyroglobulin (Tg), the only protein with a capacity to store available iodine, is a natural focus of research on iodine-accelerated SAT (ISAT) for several reasons. First, the titers of Tg-specific IgG1 and IgG2b Abs correlate with the ISAT severity,2,5 suggesting that Tg is an early autoimmune target in the disease cascade. Second, the iodine content of Tg has been shown to correlate with its immunopathogenicity in studies on Tg-induced experimental autoimmune thyroiditis (EAT),11-13 and with its antigenicity in studies of EAT14,15 and ISAT.16 Third, we have previously provided evidence that iodination may alter the immunodominance hierarchy of pathogenic T-cell epitopes within Tg,12 and that iodotyrosyl formation within certain Tg peptides may create thyroiditogenic T cell epitopes.17 Lastly, formation of Tg with a high iodine content (I-Tg) has been observed in CS chickens placed on iodine-rich drinking water,13 although this has not been confirmed in other species. In fact, we have recently demonstrated that consumption of increased amounts of iodine precipitates hypothyroidism in SJL mice with no apparent autoimmune basis.18
In the present study, we examined whether the genetic background of NOD.H2h4 mice favors formation of I-Tg when the animals follow an iodine-rich diet. We have also tested whether NOD.H2h4 mice developing ISAT generate IgG responses that preferentially recognize I-Tg vs Tg, a finding that would indirectly support formation of I-Tg in the disease process. Lastly, we have tested T-cell responses of NOD.H2h4 mice with ISAT against Tg, I-Tg and a panel of known Ak-restricted, iodinated and non-iodinated peptides of Tg, most of which are pathogenic.17,19-21 Our results do not corroborate the view that ISAT in NOD-H2h4 mice is associated with I-Tg formation or a response to neoantigenic Tg determinants containing iodine. Instead, they suggest that iodine may accelerate disease by amplifying dysregulation at other levels of the immune response.
MATERIALS AND METHODS
Mice
NOD.H2h4 mice, originally derived by Dr. L.Wicker (Merck Laboratories, Rahway, NJ), were kindly provided to us by Dr. H. Braley-Mullen (University of Missouri, Columbia, MO). The mice were bred and maintained under specific pathogen-free conditions in the animal facility of the Faculty of Medicine, Memorial University of Newfoundland. Both male and female mice were used and they were age- and sex-matched in each experiment. Mice received 0.05% NaI in their drinking water beginning at 6-8 weeks of age and thyroids or sera were obtained at various time intervals. All experimental procedures were reviewed and approved by the Animal Care Committee at Memorial University of Newfoundland.
Tg iodination and determination of I content in Tg
Frozen thyroids of BALB/c mice (Bioproducts for Science, Indianapolis, IN) were homogenized in PBS buffer, pH=7 containing 10-6 M leupeptin, 10-6 M pepstatin and 10-3 M phenylmethanesulfonylfluoride (Sigma). Thyroid extracts were centrifuged 3x at 14,000 rpm for 10 minutes at 4°C, and subsequently 920 µl samples were passed through a 1.5 cm x 47 cm Sepharose CL-4B column (Amersham Biosciences, Sweden). Tg iodination was conducted using the Iodo-Beads Iodination Reagent (Pierce, Rockford, IL) according to the manufacturer’s instructions. Briefly, one bead was added to aliquots containing 200 μl PBS and 20 μl of 2.5 mM NaI for 5 min, followed by the addition of 200 μg of Tg for 30 seconds. The beads were then removed and the samples were pooled and dialyzed 3X in water. This protocol generated mildly iodinated I-Tg, containing approximately 24 I atoms/ monomeric subunit vs 14 I atoms in control Tg. A non-incinerative method based on the catalytic activity of iodine in the ceric-arsenite reaction was used to determine Tg iodine levels, as previously described.12 The reduction of Ce(IV) to Ce(III) by As leads to a conversion of the yellow Ce ion to the colorless cerious ion, a reaction that is followed spectrophotometrically at λ=410 nm.22,23 Standard curves were constructed using known concentrations of thyroxine (T4) (Sigma) dissolved in 99 vol methanol and 1 vol 30% ammonium hydroxide. The iodine content of each Tg sample was extrapolated from the standard curve and the data are reported as the average value of two to three independent measurements ± standard deviation. Non-iodinated Tg peptides (Sigma-Genosys, The Woodlands, TX) and iodinated Tg peptides (Dalton Chemical Laboratories, Toronto, Ontario) were synthesized at >80% purity. F-moc-3-iodo-tyrosine, containing one iodine atom at position 3 of the phenolic ring, was used for the synthesis of iodinated peptides. All peptides were blocked with an acetyl group at the N terminus and with an amide group at the C terminus; the thiol group of internal Cys residues was blocked by acetamide.
Measurement of IgG responses by ELISA
Tg-specific serum IgG antibodies were detected by ELISA, as previously described.18 Briefly, wells of polyvinylchloride plates were coated overnight with 10 μg/ml Tg or I-Tg in carbonate buffer pH=9.6, washed with PBS and blocked with 0.1% BSA in PBS for 1 hour at room temperature. The plates were again washed and serum samples, diluted in PBS/Tween 20 containing 0.1% BSA, were added for 1 hour. An alkaline phosphatase-conjugated goat anti-mouse IgG antibody (Jackson Immunoresearch Laboratories, Inc) was used to detect the presence of bound antibodies. After 1 h incubation, the p-nitrophenyl phosphate substrate (Sigma) was added to the plates and light absorption was determined at 405 nm using a Vmax plate reader (Molecular Devices, Sunnyvale, CA). Results are expressed as mean O.D. values of triplicate wells ± S.D. after subtraction of background values from wells with no serum added. In competitive inhibition assays the binding of serum IgG (50 μl serum at 1:200 dilution per well) to Tg or I-Tg was monitored in the presence of increasing amounts of soluble inhibitor. Percent inhibition was determined by the formula: [1-(experim. O.D./max. O.D.)] *100 and was expressed as a mean of triplicate wells ± S.D.
[3H]-thymidine incorporation assay
At various time intervals following administration of NaI in the drinking water, cervical lymph node or spleen cell suspensions were prepared in HL-1 medium (Lonza Walkersville, MD, USA) supplemented with 100 U/ml penicillin/streptomycin, 2 mM glutamine and 50 μΜ 2-β-mercaptoethanol. Cells, adjusted at 4 x 106 cells/ml, were cultured in the presence of titrated amounts of Tg or I-Tg for 72 hours. Tg peptide reactivity was similarly assessed by using DMEM (Gibco, Invitrogen) containing 10% fetal bovine serum (PAA Laboratories Inc., Ontario, Canada) in a 96 hr assay. During the last eighteen hours of culture, 1 µCi [3H]-thymidine (Perkin Elmer, Boston, USA) was added into each well. The cells were then harvested on a ClassicCell Harvester (Skatron Instruments, Lier, Norway) and radioactivity was measured in a Beckman Coulter LS6500 scin tillation counter. Stimulation index is defined as counts per minute in the presence of antigen/counts per minute in the absence of antigen. The results are expressed as means of triplicate well values ± standard deviation (SD).
Histological assessment of thyroid infiltration by mononuclear cells
Thyroids were aseptically removed, fixed in 10% formalin, embedded in methacrylate and step-sectioned in 4 µm thick sections as previously described.24 Staining with hematoxylin and eosin followed and the presence of mononuclear cell infiltration was scored as follows: 0: no infiltration, 1: interstitial accumulation of cells between two or more follicles, 2: one or two foci of cells at least the size of one follicle, 3: diffuse infiltration of 10- 40% of the total area, 4: extensive infiltration of 40-80% of the total area, 5: extensive infiltration of more than 80% of the total area. Results report the highest infiltration index observed among the step sections of each gland.
Statistical analysis
Statistical comparison of SAT scores was performed by the non-parametric Mann-Whitney test, whereas differences in iodine content between Tg preparations were assessed by the non-parametric Wilcoxon matched pairs test, using GraphPad Prism 4.0 software (GraphPad Inc., San Diego, CA). The results were expressed means of 2-3 measurements ± SD.
RESULTS
The NOD.H2h4 genetic environment did not predispose to increased iodine incorporation in Tg
To investigate whether elevated iodide intake leads to enhanced iodine incorporation in Tg, six-week-old NOD.H2h4 mice (n=13) were placed on drinking water supplemented with 0.05% NaI for 4 weeks, whereas age-matched control NOD.H2h4 mice (n= 13) received normal drinking water for the same period. Tg was subsequently isolated from pooled thyroids by gel filtration. Since the mononuclear cell infiltration of thyroids in ISAT peaks after 8 weeks of iodide supplementation,2 it was reasoned that the thyroids of either group would yield sufficient amounts of Tg within the 4-week observation period. Surprisingly, it was found that the chromatographic elution profile of Tg from NOD.H2h4 mice did not show a 660 kDa soluble Tg peak, regardless of the dietary regimen used (Figure 1, A & B). In contrast, Tg extracted from the thyroids of CBA/J mice (n=19) placed on iodide-supplemented water or BALB/c mice (n=13) on normal water was eluted in a distinct peak (Figure 1 C & D). To investigate whether this serendipitous finding was likely to reflect the emerging Tg-specific autoreactivity in NOD.H2h4 hosts, we made use of NOD.Kμnull mice that share all the MHC and non-MHC genes of the NOD.H2h4 strain but lack B cells due to the deletion of the Ig gene and do not develop ISAT.25 We observed that Tg extracted from NOD.Kμnull mice (n=13) with or without iodide supplementation had a normal elution profile (Figure 1, E & F). Iodine determination assays, based on a non incinerative method and making use of thyroxine standard curves (Figure 2A), showed that Tg from NOD.Kμnull mice on NaI/water contained 10.9 ± 0.3 I atoms per monomer which was not statistically different from the I content of Tg from control NOD.Kμnull mice (9±2.7 I atoms per monomer) (Figure 2B). The derived values were in accordance with the I content of Tg from BALB/c mice on normal water (13.6±1.5 I atoms per monomer), or Tg from CBA/J mice on NaI/water (8.7±1.0 iodine atoms per monomer) (Figure 2B). These results suggested that the genetic environment of NOD.H2h4 mice is not a predisposing factor for increased iodine incorporation in Tg under conditions of elevated iodine intake that induce ISAT.
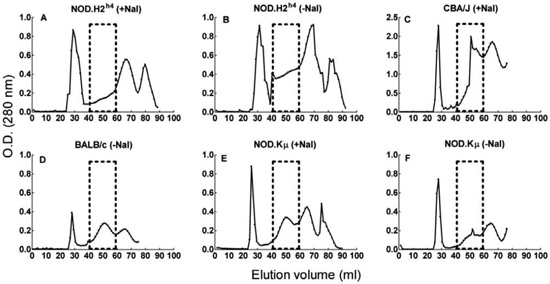
Figure 1. Elution profiles of thyroid extracts subjected to gel filtration on a Sepharose CL-4B column. Boxes indicate the elution volume of free Tg used to calibrate the column. The chromatographic elution profile of Tg from NOD. H2h4 mice did not show a 660KDa soluble Tg peak in either dietary regimen (A & B). Tg extracted from the thyroids of CBA/J mice on iodine-supplemented water or BALB/c mice on normal water was eluted in a distinct peak (C & D). Tg extracted from NOD.Kmnull mice with or without iodine supplementation had a normal elution profile (E & F).
Serum IgG responses to Tg or I-Tg preparations in ISAT
To seek immunological evidence that I-Tg formation occurs during ISAT development, we examined whether mice developing ISAT will generate IgG antibodies preferentially responding to I-Tg vs Tg. To this end, we first monitored the kinetics of ISAT development in our colony and observed a gradual increase in disease incidence with low levels of thyroid mononuclear cell infiltration 7-15 days following NaI supplementation (Table 1 ). Significant differences in both incidence and severity of ISAT were observed on d28, d42 and d77 after the initiation of NaI intake, with concomitant Tg-specific IgG responses that were significantly different between the experimental and control groups (Table 1). At a time when ISAT was well established (day 42), the serum IgG responses from iodine-fed mice did not show preferential reactivity against I-Tg vs Tg (Figure 3A). To exclude the possibility that the equivalent responses to these target molecules were not due to opposing effects of iodination on the antigenicity of Tg, i.e. blockade of prior B-cell epitopes and simultaneous formation of neoantigens by iodine atoms on Tg, we performed competition assays by ELISA. It was found that free Tg or I-Tg inhibited equivalently the IgG response to solid phase-bound Tg (Figure 3B) or I-Tg (Figure 3C). These data did not provide support for the notion that I-Tg constitutes a preferential autoimmune target in the course of ISAT development in NOD.H2h4 mice.
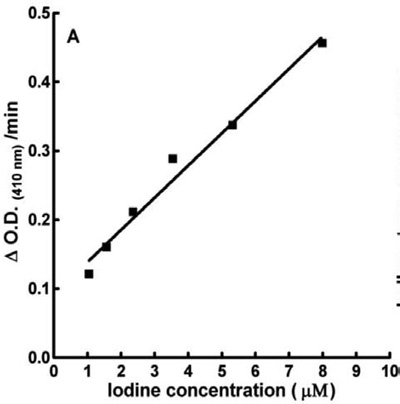
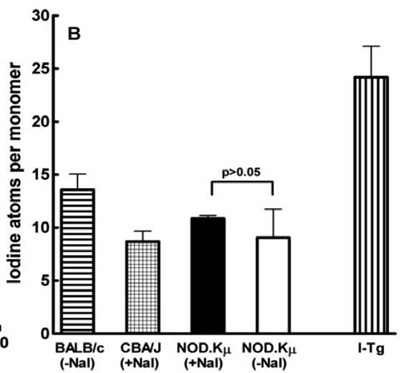
Figure 2. Determination of iodine content in Tg samples. A: Representative standard curve showing the reduction of optical density at 410 nm of the iodine-catalyzed ceric-arsenite reaction as a function of T4 added. B: Iodine content of Tg preparations isolated, as shown in Figure 1, from thyroids of mice receiving NaI/drinking water (+NaI) or controls (-NaI). Each bar represents the mean iodine content of Tg±SD from two to three independent measurements. The last bar shows the iodine content of in vitro-iodinated Tg (I-Tg) purified from BALB/c mice and used as a target Ag in ELISA and cell proliferation assays. Statistical significance was calculated by the non-parametric Wilcoxon matched pairs test. There was no significant difference (p>0.05) in the iodine content of Tg preparations isolated from NOD.Kμnull mice regardless of the dietary regimen followed.
Proliferative responses of NOD.H2h4 lymphoid cells to Tg, I-Tg and Tg peptides during ISAT development
We next examined possible differences between Tg and I-Tg antigenicity at the T cell level, in early and late stages of ISAT development. NOD.H2h4 thyroid-draining cervical lymph node cells, isolated after 7 days of iodide supplementation, were cultured in the presence of Tg or I-Tg and their proliferative responses were examined. The SI values against either antigen were generally undetectable (SI <2), providing no evidence for differential reactivity to I-Tg in early stages of ISAT (Figure 4A). After 28 or 42 days of iodide supplementation, the spleen cell responses to Tg and I-Tg were low but comparable (SI values between 2-4) (Figure 4 B, C). To investigate possible reactivity against known Tg T-cell epitopes, we used a panel of Ak-binding Tg peptides that included the iodinated epitopes I-p117, I-p179, I-p1931, I-p2529, I-p 254017,20 and the non-iodinated peptides p226, p1579, p1826 and p2026.17,21 NOD.H2h4 splenic cells, obtained 8 weeks after the initiation of NaI supplementation, showed a uniform lack of response (SI <2) to these peptides (Figure 4 D, E). Collectively, the above data did not provide evidence that ISAT is associated with T-cell responses to putative I-Tg molecules or known thyroiditogenic iodinated Tg peptides in NOD.H2h4 mice.
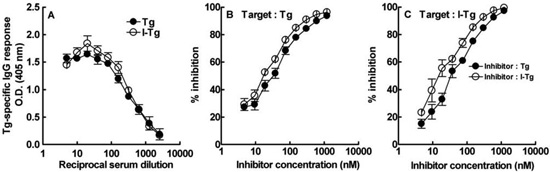
Figure 3. A: Equivalent serum IgG responses to Tg (•) or I-Tg () preparations, assayed by ELISA, at a time (day 42) when ISAT is established in NOD.H2h4 mice with high dietary iodine intake. Results are expressed as mean O.D. values of triplicate wells ± S.D; B: Competitive inhibition of IgG binding to solid phase-bound Tg. Equivalent inhibition was mediated by soluble Tg (•) or I-Tg () preparations; C: Competitive inhibition of IgG binding to solid phase-bound I-Tg suggesting that I-Tg () is not a better inhibitor than Tg (•). Initial concentration of inhibitor was 1210 nM and sera were used at 1:200 dilution. O.D. values from wells without inhibitor ranged from 1.1-1.2. The inhibition using OVA as a control competitor did not exceed 8%.
DISCUSSION
The iodine-mediated acceleration of spontaneous thyroiditis in NOD.H2h4 mice has been well confirmed in several studies2,4,5 but the reasons behind this intriguing observation remain obscure. Since Tg serves as a storage molecule of available iodine, we have examined the iodine content of Tg in mice developing ISAT, seeking evidence for I-Tg formation in vivo that might be linked to the higher incidence and greater severity of the disease. Our study was initially hindered by the fact that the 660 kDa peak of soluble, free Tg was not discernible in chromatograms of thyroid extracts from NOD.H2h4 mice, especially those exhibiting both mild ISAT (mean I.I. = 2.00) and circulating Tg-specific serum IgG, four weeks after the initiation of the iodine-rich diet. The formation of larger than usual pellets in centrifuged thyroid extract samples prior to gel filtration suggested possible generation of Tg-Ab complexes either in vivo or during the Tg purification procedure. To circumvent this problem, we purified Tg from thyroids of B-cell deficient NOD.H2h4 mice (NOD.Kμnull) which do not develop ISAT when they receive NaI in their drinking water.25 We found no significant difference in the iodine content of Tg between control and experimental groups (9.0 ± 2.7 vs 10.9 ± 0.3 I atoms per monomeric Tg subunit). Our data are in apparent contradiction with those of Sundick et al. who observed that CS chickens, which are susceptible to iodide-induced thyroiditis, incorporate at least 60 I atoms per Tg molecule when placed on a high iodide diet (20 mg KI/dl drinking water).13 The reasons behind this discrepancy are unclear, and the extensive immune dysregulation contributed by non-MHC genes in the NOD background26 precludes meaningful comparisons between the two models.
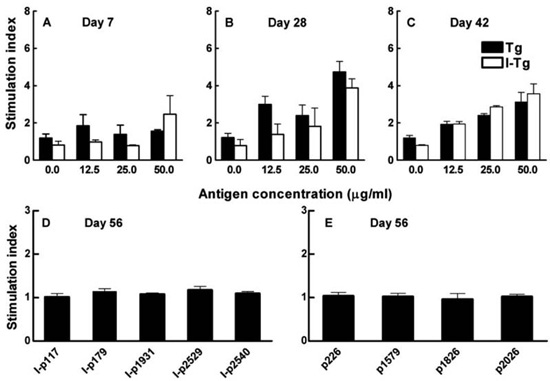
Figure 4. Proliferative responses to Tg and I-Tg using NOD.H2h4 cervical lymph node cells (A) or splenic cells (B &C) (two mice per group), on the indicated days following initiation of iodide supplementation. Background levels were: A: 500 cpm, B: 1800 cpm and C: 760 cpm. Proliferative responses to a panel of iodinated (D) and non-iodinated (E) Ak –binding Tg peptides at 10 μM using splenocytes from male NOD.H2h4 mice with ISAT. Background levels were 8800 cpm. Results are expressed as means of triplicate well values±SD. The overall data do not support the view that, after iodination, intact Tg or Tg peptides become preferential targets of autoreactive T cells.
The fact that Tg-specific Abs are readily detected in ISAT2,4,5 prompted us to look for putative differences in B-cell reactivity against Tg vs I-Tg preparations. We hypothesized that chronic iodide consumption might lead to intrathyroidal generation of Tg with slightly higher than normal iodine content that might become a preferred target in the autoimmune process. Thus, I-Tg was prepared in vitro with approx. 24 I atoms per monomer and its antigenic profile was compared to that of normal Tg in ELISA. After 7 weeks of iodide supplementation, i.e. at a time when ISAT is well developed, the serum IgG responses were equally strong against I-Tg and Tg. Competitive inhibition assays provided no evidence that iodide in I-Tg blocked existing determinants or facilitated formation of neoantigenic epitopes in a manner that could skew the autoantibody response. These data did not provide indirect support for I-Tg generation in ISAT, although it cannot be formally excluded that Tg iodination in vitro may differ both qualitatively and quantitatively from the enzyme-mediated intrathyroidal iodination process.
The ease of detection of Tg-specific Abs in ISAT is in apparent contrast with the very low level of T-cell reactivity to Tg observed either in thyroid draining lymph nodes or spleens of NOD.H2h4 mice.2,27 Our results confirm this finding, which is reminiscent of analogous observations regarding the insulin-specific responses in NOD mice developing spontaneous diabetes.26,28 It has been reported that the natural autoantibody repertoire of NOD mice is highly active against self-antigens such as insulin, Tg and DNA29 and it is possible that genetic abnormalities in the regulation of Tg-reactive B cells in NOD.H2h4 mice may contribute to autoimmunity. Iodide has also been reported to enhance IgG synthesis by human peripheral blood lymphocytes in vitro.30 In our study, the detection of low but equivalent proliferative cell responses to Tg and I-Tg in lymphoid organs of NOD.H2h4 mice developing ISAT additionally suggested that the iodine content of Tg does not affect its antigenicity in this model. Furthermore, we could not detect proliferative cell responses of NOD.H2h4 spleen cells to Ak-restricted T cell epitopes of Tg most of which are known to induce thyroiditis either as iodinated17,20 or non-iodinated peptides.17,21
Our inability to demonstrate formation of I-Tg in this ISAT model is seemingly at odds with the results of numerous EAT studies6,11,19,31,32 attributing a critical role for Tg iodination in the disease process. In several studies, iodine-depleted Tg does not activate thyroiditogenic T cells but, by raising the Tg iodine content to normal levels in vitro, the immunopathogenicity of the molecule is re-established.11,31 At the other end of the spectrum, highly iodinated Tg may cause EAT with increased incidence and severity in SJL mice,12 but when these mice are fed a high iodine diet, their Tg preparations do not show high iodine incorporation ex vivo and the mice develop goitrous hypothyroidism with no apparent autoimmune basis.18 To our knowledge, in vivo formation of I-Tg in normal (i.e. non-goitrous) mice exposed to a chronic iodine-rich diet has not yet been reported.
If enhanced iodine intake does not cause ISAT via an I-Tg “link” in NOD.H2h4, mice what might be its mode of action? In addressing this question, it is worth mentioning that in the ISAT model a mouse consuming on average 5 ml of 0.05% NaI/water daily, receives approximately 2 mg of iodide per day, i.e. a 2000-fold higher amount than that present in normal diet.33 This dietary regimen has recently been shown to cause damage in the ultrastructure of NOD.H2h4 thyroid epithelial cells associated with apoptosis and necrosis,34 a process that might be influenced by production of high H2O2 levels in NOD.H2h4 thyrocytes.35 In addition, enhanced iodine intake may upregulate adhesion molecules such as ICAM-1 on thyrocytes,36 thus accelerating mononuclear cell infiltration into the thyroid. This process has been postulated to ope¬rate via the elevated generation of reactive oxygen intermediates.35,37 However, the same dietary regimen does not appear to cause overt histological changes in the thyrocytes of other mouse strains – except for SJL18 – and the genetic factors predisposing NOD.H2h4 mice to ISAT remain unknown. Finally, iodine may amplify inherent defects in immune cells involved in the autoimmune response, such as dendritic cells, which have been reported to exhibit phenotypic and functional abnormalities in NOD.H2h4 mice as compared to dendritic cells from CBA mice.38 Iodine may also directly impair the function of regulatory T cells that have been known to contribute to ISAT resistance in NOD.H2h4 mice.39-41 Future research efforts in this area may yield new insights into the pathogenesis of thyroiditis.
ACKNOWLEDGMENTS
We thank Dr. H. Braley-Mullen for the NOD.H2h4 breeding pairs and her kind provision of thyroids from NOD.Kμnull mice used in this study.
REFERENCES
1. Podolin PL, Pressey A, DeLarato NH, Fischer PA, Peterson LB, Wicker LS, 1993 I-E+ nonobese diabetic mice develop insulitis and diabetes. J Exp Med 178: 793-803.
2. Braley-Mullen H, Sharp GC, Medling B, Tang H, 1999 Spontaneous autoimmune thyroiditis in NOD.H-2h4 mice. J Autoimmun 12: 157-165.
3. Burek CL, Sharma RB, Rose NR, 2003 NKT cell regulation of autoimmune thyroiditis. Autoimmunity 36: 405-408.
4. Hutchings PR, Verma S, Phillips JM, Harach SZ, Howlett S, Cooke A, 1999 Both CD4(+) T cells and CD8(+) T cells are required for iodine accelerated thyroiditis in NOD mice. Cell Immunol 192: 113-121.
5. Rasooly L, Burek CL, Rose NR, 1996 Iodine-induced autoimmune thyroiditis in NOD-H-2h4 mice. Clin Immunol Immunopathol 81: 287-292.
6. Bagchi N, Brown TR, Urdanivia E, Sundick RS, 1985 Induction of autoimmune thyroiditis in chickens by dietary iodine. Science 230: 325-327.
7. Allen EM, Appel MC, Braverman LE, 1986 The effect of iodide ingestion on the development of spontaneous lymphocytic thyroiditis in the diabetes-prone BB/W rat. Endocrinology 118: 1977-1981.
8. Bournaud C, Orgiazzi JJ, 2003 Iodine excess and thyroid autoimmunity. J Endocrinol Invest 26: 49-56.
9. Konno N, Makita H, Yuri K, Iizuka N, Kawasaki K, 1994 Association between dietary iodine intake and prevalence of subclinical hypothyroidism in the coastal regions of Japan. J Clin Endocrinol Metab 78: 393-397.
10. Markou K, Georgopoulos N, Kyriazopoulou V, Vagenakis AG, 2001 Iodine-Induced hypothyroidism. Thyroid 11: 501-510.
11. Champion BR, Rayner DC, Byfield PG, Page KR, Chan CT, Roitt IM, 1987 Critical role of iodination for T cell recognition of thyroglobulin in experimental murine thyroid autoimmunity. J Immunol 139: 3665-3670.
12. Dai YD, Rao VP, Carayanniotis G, 2002 Enhanced iodination of thyroglobulin facilitates processing and presentation of a cryptic pathogenic peptide. J Immunol 168: 5907-5911.
13. Sundick RS, Herdegen DM, Brown TR, Bagchi N, 1987 The incorporation of dietary iodine into thyroglobulin increases its immunogenicity. Endocrinology 120: 2078-2084.
14. Saboori AM, Rose NR, Burek CL, 1998 Iodination of human thyroglobulin (Tg) alters its immunoreactivity. II. Fine specificity of a monoclonal antibody that recognizes iodinated Tg. Clin Exp Immunol 113: 303-308.
15. Saboori AM, Rose NR, Bresler HS, Vladut-Talor M, Burek CL, 1998 Iodination of human thyroglobulin (Tg) alters its immunoreactivity. I. Iodination alters multiple epitopes of human Tg. Clin Exp Immunol 113: 297-302.
16. Barin JG, Talor MV, Sharma RB, Rose NR, Burek CL, 2005 Iodination of murine thyroglobulin enhances autoimmune reactivity in the NOD.H2 mouse. Clin Exp Immunol 142: 251-259.
17. Li HS, Carayanniotis G, 2006 Iodination of tyrosyls in thyroglobulin generates neoantigenic determinants that cause thyroiditis. J Immunol 176: 4479-4483.
18. Li HS, Carayanniotis G, 2007 Induction of goitrous hypothyroidism by dietary iodide in SJL mice. Endocrinology 148: 2747-2752.
19. Carayanniotis G, 2007 Recognition of thyroglobulin by T cells: the role of iodine. Thyroid 17: 963-973.
20. Li HS, Jiang HY, Carayanniotis G, 2007 Modifying effects of iodine on the immunogenicity of thyroglobulin peptides. J Autoimmun 28: 171-176.
21. Verginis P, Stanford MM, Carayanniotis G, 2002 Delineation of five thyroglobulin T cell epitopes with pathogenic potential in experimental autoimmune thyroiditis. J Immunol 169: 5332-5337.
22. Palumbo G, Tecce MF, Ambrosio G, 1982 A nonincinerative rate-sensing method for the determination of iodine in iodoproteins. Anal Biochem 123: 183-189.
23. Saboori AM, Rose NR, Butscher WG, Burek CL, 1993 Modification of a nonincinerative method for determination of iodine in iodoproteins. Anal Biochem 214: 335-338.
24. Rao VP, Kajon AE, Spindler KR, Carayanniotis G, 1999 Involvement of epitope mimicry in potentiation but not initiation of autoimmune disease. J Immunol 162: 5888-5893.
25. Braley-Mullen H, Yu S, 2000 Early requirement for B cells for development of spontaneous autoimmune thyroiditis in NOD.H-2h4 mice. J Immunol 165: 7262-7269.
26. Anderson MS, Bluestone JA, 2005 The NOD mouse: a model of immune dysregulation. Annu Rev Immunol 23: 447-485.
27. Verma S, Hutchings P, Guo J, McLachlan S, Rapoport B, Cooke A, 2000 Role of MHC class I expression and CD8(+) T cells in the evolution of iodine-induced thyroiditis in NOD-H2(h4) and NOD mice. Eur J Immunol 30: 1191-1202.
28. Kaufman DL, Tisch R, Sarvetnick N et al, 2001 Report from the 1st International NOD Mouse T-Cell Workshop and the follow-up mini-workshop. Diabetes 50: 2459-2463.
29. Thomas JW, Kendall PL, Mitchell HG, 2002 The natural autoantibody repertoire of nonobese diabetic mice is highly active. J Immunol 169: 6617-6624.
30. Weetman AP, McGregor AM, Campbell H, Lazarus JH, Ibbertson HK, Hall R, 1983 Iodide enhances IgG synthesis by human peripheral blood lymphocytes in vitro. Acta Endocrinol (Copenh) 103: 210-215.
31. Ebner SA, Lueprasitsakul W, Alex S, Fang SL, Appel MC, Braverman LE, 1992 Iodine content of rat thyroglobulin affects its antigenicity in inducing lymphocytic thyroiditis in the BB/Wor rat. Autoimmunity 13: 209-214.
32. Ruwhof C, Drexhage HA, 2001 Iodine and thyroid autoimmune disease in animal models. Thyroid 11: 427-436.
33. Rose NR, Bonita R, Burek CL, 2002 Iodine: an environmental trigger of thyroiditis. Autoimmun Rev 1: 97-103.
34. Teng X, Shan Z, Teng W, Fan C, Wang H, Guo R, 2009 Experimental study on the effects of chronic iodine excess on thyroid function, structure, and autoimmunity in autoimmune-prone NOD.H-2h4 mice. Clin Exp Med 9: 51-59.
35. Burek CL, Rose NR, 2008 Autoimmune thyroiditis and ROS. Autoimmun Rev 7: 530-537.
36. Bonita RE, Rose NR, Rasooly L, Caturegli P, Burek CL, 2002 Adhesion molecules as susceptibility factors in spontaneous autoimmune thyroiditis in the NOD-H2h4 mouse. Exp Mol Pathol 73: 155-163.
37. Sharma R, Traore K, Trush MA, Rose NR, Burek CL, 2008 Intracellular adhesion molecule-1 up-regulation on thyrocytes by iodine of non-obese diabetic.H2(h4) mice is reactive oxygen species-dependent. Clin Exp Immunol 152: 13-20.
38. Strid J, Lopes L, Marcinkiewicz J et al, 2001 A defect in bone marrow derived dendritic cell maturation in the nonobesediabetic mouse. Clin Exp Immunol 123: 375-381.
39. Nagayama Y, Horie I, Saitoh O, Nakahara M, Abiru N, 2007 CD4+CD25+ naturally occurring regulatory T cells and not lymphopenia play a role in the pathogenesis of iodide-induced autoimmune thyroiditis in NOD-H2h4 mice. J Autoimmun 29: 195-202.
40. Nakahara M, Nagayama Y, Saitoh O, Sogawa R, Tone S, Abiru N, 2008 Expression of Immuno-Regulatory Molecules by Thyrocytes Protects NOD-H2h4 Mice from Developing Autoimmune Thyroiditis. Endocrinology 150: 1545-1551
41. Yu S, Maiti PK, Dyson M, Jain R, Braley-Mullen H, 2006 B cell-deficient NOD.H-2h4 mice have CD4+CD25+ T regulatory cells that inhibit the development of spontaneous autoimmune thyroiditis. J Exp Med 203: 349-358.
Address for correspondence:
George Carayanniotis, Faculty of Medicine, Memorial University of Newfoundland, St. John’s NL, Canada A1B 3V6,
Tel: 001-709-777- 6967, Fax: 001-709-777-7010, e-mail: gcarayan@mun.ca
Received 24-11-09, Revised 08-02-10, Accepted 25-02-10